МОРФОДИНАМИКА
ОДНОЛЕТНИХ ПОБЕГОВ ИВЫ ШЕРСТИСТОПОБЕГОВОЙ
И
ПЕРСПЕКТИВЫ ИСПОЛЬЗОВАНИЯ КОНЦЕПЦИИ МОРФОДИНАМИЧЕСКИХ АТТРАКТОРОВ ДЛЯ ОЦЕНКИ
ГЕНЕТИЧЕСКОГО ПОТЕНЦИАЛА ПРИРОДНЫХ ПОПУЛЯЦИЙ
Афонин Алексей Алексеевич
– д. с.-х. н., проф. кафедры зоологии и анатомии Брянского государственного
университета
Анищенко Лидия
Николаевна – д. с.-х. н., доц. кафедры экологии и рационального
природопользования Брянского государственного университета
Печатный
вариант:
Афонин
А.А. Морфодинамика однолетних побегов ивы шерстистопобеговой и перспективы
использования концепции морфодинамических аттракторов для оценки генетического
потенциала природных популяций / А.А. Афонин, Л.Н. Анищенко // Стратегия
устойчивого развития регионов России: Сб. материалов III Всеросс.
научно-практ. конфер. – Новосибирск: Изд-во НГТУ,
2010. – С. 125–132.
Ива
шерстистопобеговая (Salix dasyclados Wimm.)
– перспективный вид для создания высокопродуктивных энергетических плантаций.
Для оценки генетического потенциала природных популяций проведен сравнительный
анализ метамерной изменчивости ширины листовой пластинки. Впервые для решения
подобных задач использованы параметрически заданные XX'-кривые.
Выявлены морфодинамические аттракторы в виде петель на фазовой плоскости X–X', что позволило
установить различия в морфодинамике побегов. Раскрыты
перспективы создания квазинативных устойчивых поликлональных насаждений.
Угроза
истощения невозобновляемых энергетических ресурсов
требует поиска новых («альтернативных») аккумуляторов лучистой энергии Солнца –
основного условно возобновляемого источника энергии на Земле. Среди множества
решений этой проблемы видное место занимает биоэнергетика – раздел энергетики,
основанный на биоаккумуляции солнечной энергии и
дальнейшем переработке полученной биомассы для хранения и преобразования
полученной энергии. Биомасса быстрорастущих деревьев и кустарников («биотопливо
второго поколения») представляет собой один из самых дешевых и безопасных биоаккумуляторов [Цивенкова, Самылин, 2005]. Древесно-кустарниковые биоэнергетические плантации могут
создаваться на малоплодородных неокультуренных почвах (непригодных для
возделывания большинства сельскохозяйственных растений), что снимает
противоречие между энергетической и продовольственной проблемами.
Поэтому в странах, бедных традиционными энергоносителями, вопросам, связанным с
проектированием биоэнергетических плантации, уделяется все больше внимания
[Егоров, 2005; McCracken,
1998].
В
качестве источника биомассы широко используются два вида кустарниковых ив:
трехтычинковая (Salix
triandra L.) и корзиночная
(S.
viminalis L.), а также их гибриды [Bollmark et al., 1999; Dawson, McCracken, 1998]. Однако эти
виды могут нормально произрастать только на хорошо дренированных почвах. В
результате создание ивовых плантаций на тяжелых переувлажненных почвах
оказывается невозможным. Поэтому необходимо искать такие высокопродуктивные
виды ив, которые могли бы возделываться в условиях, непригодных для S. triandra
и S.
viminalis.
Одним
из таких видов является ива шерстистопобеговая (Salix dasyclados
Wimm.). Это крупный быстрорастущий
геоксильный кустарник, реже дерево. Легко черенкуется. Порослевые побеги
прутьевидные, очень мощные. В естественном состоянии произрастает на сырых почвах,
в понижениях центральной и прирусловой поймы, вне поймы обычна
на сырых лугах, вдоль дорог; отсутствует на болотах с избыточно застойным
увлажнением [Валягина-Малютина, 2004].
Ареал
евроазиатского бореального типа: Центральная и Северная Европа, Сибирь, Дальний
Восток, Монголия, Китай, Япония. Вследствие географической подразделенности видового ареала описаны географические
расы, которым М.И. Назаровым [Назаров, 1936] был присвоен ранг вида: S.
burjatica Nas., S. jacutica Nas. В то же время
внутрипопуляционная (формовая) изменчивость ивы шерстистопобеговой изучена
недостаточно. Не исключено, что изучению формового разнообразия препятствует
высокий уровень полиплоидии (2n=114
[Числа хромосом…, 1993]; возможно, S.
dasyclados
является
гибридом S. cinerea
× S. viminalis [Цвелев, 2000]),
что приводит к забуференности популяционных
генофондов. Полиплоидность ивы шерстистопобеговой
проявляется в том, что она отличается от других ив средней полосы России
наиболее крупными пыльцевыми зернами: длина полярной оси 28,9…37,3 (31,7…34,8)
мкм [Афонин, 2006]. Это позволяет отличать тычиночные особи ивы
шерстистопобеговой от других видов со сходной морфологией побегов.
Затруднения
в оценке уровня внутрипопуляционного разнообразия ивы шерстистопобеговой по
морфологическим признакам являются препятствием для выделения
высокопродуктивных клонов, различающихся по ритмам развития побегов. Коллекции
таких клонов могут использоваться для создания полифункциональных насаждений (в
т.ч., и энергетических плантаций), в которых наиболее полно реализуется
суммарный информационно-энергетический потенциал. Эти плантации, по существу,
должны представлять собой генетически гетерогенные, эволюционно устойчивые
квазинативные экосистемы с максимально возможным уровнем биологического разнообразия
и оптимизированным уровнем конкуренции [Афонин, 2009с].
На
основании многолетних наблюдений за изменчивостью ивы шерстистопобеговой на
территории Брянского лесного массива [Афонин, 2002, 2003] нами было
установлено, что типичные экземпляры этого вида характеризуются широко ланцетовидными листьями длиной 70...105 (40...140) мм
при ширине 20...30 (15...35) мм. Показатели средней внутрикронной и межиндивидуальной вариабельности параметров листовой
пластинки приведены в таблице (Х –
ширина листовой пластинки, Y – длина листовой пластинки, М – среднее значение каждого параметра
листа, СВК
– средний внутрикронный коэффициент изменчивости, СМИ – межиндивидуальный
коэффициент изменчивости).
Таблица – Вариабельность
ширины и длины листовой пластинки ивы шерстистопобеговой
|
Статистические показатели |
Параметры листовой пластинки |
|
|
X |
Y |
|
|
М, мм |
24,5 |
87,0 |
|
С ВК, % |
21,7 |
30,0 |
|
С МИ, % |
10,9 |
16,1 |
Низкая
межиндивидуальная вариабельность ширины листовой пластинки потребовала новых
подходов к выявлению закономерностей внутрипопуляционной изменчивости данного
параметра. В настоящей работе предпринята попытка выявить группы побегов,
различающиеся по ритмам развития, а именно, по метамерной изменчивости ширины
листовой пластинки.
Еще
в 1917 г. Д’Арси Томпсон, основываясь на работах Гофмейстера и Бозе, обратил
внимание на циклический характер развития растений [Магомедмирзаев, 1990]. В
нашей стране первопроходцем в области изучения онтогенетической изменчивости
был, несомненно, Н.П. Кренке [Кренке, 1940], проанализировавший одновершинные и
двухвершинные возрастные кривые, отражающие жизнеспособность и мощность
последовательно образующихся органов. К настоящему времени описаны
одновершинные кривые изменения длины междоузлия вдоль побега для ивы
трехтычинковой [Михалевская, 1995], а также поузловая изменчивость размеров
почек и листьев ивы розмаринолистной [Гетманец, 1994].
Нами
детально описана метамерная изменчивость побегов ив пепельной
и трехтычинковой [Афонин, 2000, 2009а; 2009b; Афонин, Самошкин, 2006]. Однако
метамерная изменчивость побегов ивы шерстистопобеговой до сих пор изучена
недостаточно.
Материал
для исследования собирался 8…14 июля 2010 г. (в конце раннелетнего периода
развития) в центральной пойме р. Десны (на территории Снежетьско-Деснянского низменного (долинно-речного)
ландшафта) в зеленой зоне г. Брянска, в пределах ассоциации Salicetum triandro-viminalis (Tx. 1931) Lohm. 1952,
визуально ограниченной координатами 34,371874…34,366343º в.д. и
53,220569…53,224202º с.ш. Для получения исходных данных по побегам с
каждой особи, обладающей типичными признаками вида,
срезались завершившие рост неразветвленные ростовые побеги (ауксибласты). С
одной особи срезалось 10 побегов. Всего описано 18 особей (выполнено 3420
измерений). После отсева особей с явными повреждениями или нарушениями
морфогенеза побегов для окончательного анализа были оставлены особи 107, 109,
110, 111, 112.
За
структурно-функциональную единицу побега был принят метамер, который
рассматривался как узел с листом, пазушной почкой и нижележащим междоузлием
[Магомедмирзаев, 1990]. Для каждого метамера с порядковым номером t определялась ширина
листовой пластинки Х с точностью ±0,5 мм. Для создания баз данных
и обработки результатов использовался процессор Excel.
Метамерная
(порядковая, поузловая) изменчивость ширины листовой пластинки от порядкового
номера метамера (узла) t
описывалась с помощью морфодинамических рядов, графическим отражением
которых являются морфодинамические кривые. Прилагательное
«морфодинамический» использовалось ранее для обозначения особых точек
(морфодинамических центров), концентрирующих энергию и воздействующих на
вещество, собирающих его вокруг себя и становящихся видимыми и морфологически
наблюдаемыми [Магомедмирзаев, 1990]. Мы вкладываем в этот термин иной смысл, и
рассматриваем морфодинамический ряд (и его графическое отражение –
морфодинамическую кривую) как ось креода
– устойчивую информационно-энергетическую траекторию онтогенеза (морфогенеза),
сформировавшуюся в результате канализирующего отбора.
Визуально
однородные эмпирические кривые побегов объединялись в группы; в пределах одной
особи насчитывалось до 4 групп побегов. Полученные группы морфодинамических
кривых рассматривались как реализации случайной функции:
X (t) = φ (t) + U (t),
где φ (t) – генетически детерминированные
математические ожидания Х при заданных значениях порядкового
номера метамера t,
а U(t) – случайные величины, зависящие
от комплекса парагенетических факторов [Гмурман, 1999].
Для
каждой группы побегов вычислялось среднее значение Х для каждого t. Учитывая,
что полученные временные ряды являются нечеткими, для каждого ряда применялось
выравнивание методом скользящей средней по пяти смежным точкам t без взвешивания.
Для
аппроксимации математических ожиданий случайных функций использовались полиномы
5-й степени:
X = a5·t5
+ a4·t4 + a3·t3
+ a2·t2 + a1·t + a0,
где t –
порядковый номер метамера, a
–
эмпирические коэффициенты. Количественная адекватность уравнений регрессии
проверялась с помощью F-критерия
Фишера [Львовский, 1982]. Качественную адекватность
полученных уравнений определяли с помощью коэффициентов аппроксимации R2.
Полученные
графики представляют собой вычисленные (аппроксимированные) морфодинамические
кривые (α-кривые), отражающие основные закономерности метамерной
изменчивости ширины листовой пластинки.
Всего
было проанализировано 14 α-кривых. При этом было установлено, что
некоторые α-кривые побегов, сходных по мощности
развития, пересекаются в особых областях с минимальной дисперсией Х
при заданном t. Область пересечения
трех и более α-кривых представляет собой
морфодинамический аттрактор, прохождение через который обеспечивает полную
реализацию информационно-энергетического потенциала группы побегов. Пример
такого аттрактора показан на рисунке 1 (в легенде приведены номера особей,
через дефис указаны группы побегов).
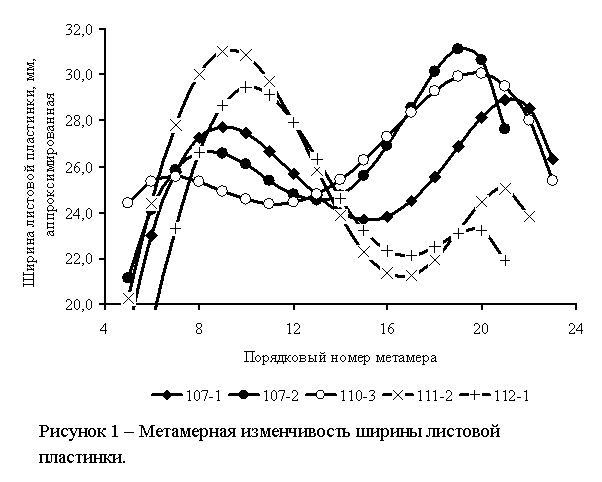
Однако
предложенное определение аттрактора не соответствует общепринятым
представлениям об аттракторах как особых точках (циклах) в фазовых плоскостях
[Свирижев, Пасеков, 1982]. Поэтому нами была предпринята попытка анализа изменчивости ширины листовой пластинки ивы
шерстистопобеговой в фазовой плоскости X–X',
где X'
– первая производная полинома 5-й степени:
X' =
5a5·t4 + 4a4·t3
+ 3a3·t2 + 2a2·t
+ a1.
Фазовые
портреты групп побегов показаны на рисунке 2.
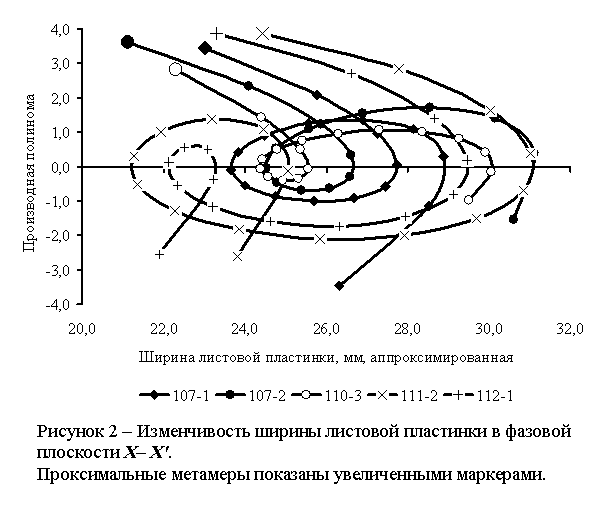
Сравнительный
анализ полученных параметрически заданных XX'-кривых показал
следующее.
1)
Все XX'-кривые
являются самопересекающимися. XX'-кривые
111-2 и 112-1 пересекают ось Х дважды, а XX'-кривые 107-1,
107-2 и 110-3 – трижды, причем определенные участки XX'-кривых образуют
петли с выходом из цикла.
2)
Петли с выходом из цикла представляют собой морфодинамические аттракторы. Выход
из цикла отличает их от физических аттракторов – замкнутых циклов или
сходящихся спиралей. Таким образом, метастабильное состояние побега (открытой
термодинамической системы) принципиально отличается от метастабильного
состояния в закрытых физико-химических системах.
3)
Центры аттракторов лежат вблизи точек с координатами X'=0, однако
смещены по оси Х. Кроме того, аттракторы различаются по пределам
изменчивости Х при X'=0.
Это позволяет выделить две группы аттракторов: [107-1, 107-2, 110-3] с центром Х=25,0…25,7
мм и [111-2, 112-1] с центром Х=22,7…23,2 мм.
4)
Петля 107-2 лежит внутри петли 107-1. Таким образом, несмотря на явные различия
между α-кривыми этих групп побегов одной особи,
можно утверждать, что закономерности морфодинамики побегов определяются
индивидуальными особенностями особи.
Полученные
результаты позволяют утверждать, что внешне сходные особи ивы
шерстистопобеговой различаются по ритмам развития побегов, в частности, по
метамерной изменчивости ширины листовой пластинки. Это позволяет рационально
использовать генетический потенциал природных популяций ивы шерстистопобеговой,
а именно: вести целенаправленный отбор комплементарных
дендроритмотипов, которые могут быть использованы для
создания квазинативных генетически гетерогенных высокопродуктивных насаждений,
максимально использующих ресурсы среды и способных противостоять
неблагоприятному воздействию разнообразных стрессоров [Афонин, 2009b, 2009с].
Список литературы
1. Афонин
А.А. Ивы Брянского лесного массива. – В 2-х частях. – Брянск: БГУ, 2003. – 237
с.
2. Афонин
А.А. Изменчивость длины пыльцевых зерен аборигенных видов ив
Брянского лесного массива // ИВУЗ. Лесн. журн. – 2006. – № 5. – С.
25–34.
3. Афонин
А.А. Метамерная изменчивость листьев ивы пепельной // Наука и образование –
возрождению сельского хозяйства России в XXI веке: Междунар. науч.-практ. и учебно-метод. конф.
– Брянск: Брянская гос. с.-х. академия, 2000. – С.
70–72.
4. Афонин
А.А. Метамерная изменчивость листьев ивы трехтычинковой / А.А. Афонин, Е.Н.
Самошкин // Лесоведение. – 2006. – № 2. – С. 5–10.
5. Афонин
А.А. Методы анализа морфодинамики однолетних побегов ив (Salix) – [Электрон.
ресурс]. – 2009а / Код доступа – http://afonin-59-salix.narod.ru/060825_dynamica.htm
6. Афонин
А.А. Методы изучения разнообразия ив в коллекционных насаждениях. – Брянск:
Брянский гос. ун-т, 2002. – 67 с.
7. Афонин
А.А. Селекция ив на разнообразие ритмов развития // Фактори експериментальноï еволюцiï организмiв: збiрник
наукових праць (Материалы V междунар. научн. конфер.
«Факторы экспериментальной эволюции организмов»). –
Т.6. – Киïв:
Логос, 2009b.
– С. 259–263.
8. Афонин
А.А. Сохранение и приумножение уровня биологического
разнообразя в квазинативных экосистемах / А.А. Афонин, Л.И. Булавинцева
/ Биологические эффекты малых доз ионизирующей радиации и радиоактивное
загрязнение среды: Материалы Междунар. конф. – Сыктывкар: Коми НЦ УрО РАН, 2009с. – С. 291–393.
9. Валягина-Малютина
Е.Т. Ивы европейской части России. – М.: КМК, 2004. –
217 с.
10. Гетманец,
И.А. Морфологические особенности побегов и жизненные формы Salix rosmarinifolia
L.,
Salix
repens L. // Успехи экологической
морфологии растений и ее влияние на смежные науки. М.: Изд-во «Прометей» МПГУ, 1994. – С. 20-21.
11. Гмурман
В.Е. Теория вероятностей и математическая статистика. – М.: Высш. шк., 1999. –
479 с.
12. Егоров
А.И. Ивовая энергетика [Электрон. ресурс].
– 2005 / Код доступа – http://hepd.pnpi.spb.ru/ioc/ioc/line051112/n5.htm
13. Кренке
Н.П. Теория циклического старения и омоложения растений. – М.: Огиз–Сельхозгиз, 1940. – С.
5–120.
14. Львовский,
Е.Н. Статистические методы построения эмпирических формул. – М.: Высш. шк.,
1982. – 224 с.
15. Магомедмирзаев
М.М. Введение в количественную морфогенетику. – М.:
Наука, 1990. – 229 с.
16. Михалевская
О.Б. Структура и развитие боковых силлептических побегов у древесных растений и
представление об элементарном побеге / Актуальные вопросы экологической
морфологии растений. – М.: Прометей, 1995. – С. 70–76.
17. Назаров
М.И. Salix
/ Флора СССР. – Т. 5. – М.–Л.: Изд-во АН СССР, 1936. – С. 24-216, 707-713.
18. Свирижев
Ю.М. Основы математической генетики / Ю.М. Свирижев, В.П. Пасеков // М.: Наука,
1982. – 512 с.
19. Цвелев
Н.Н. Определитель сосудистых растений Северо-Западной России (Ленинградская,
Псковская и Новгородская области). – СПб.: Изд-во СПХФА,
2000. – 781 с.
20. Цивенкова
Н.М. Леса – неисчерпаемые энергоресурсы России / Н.М. Цивенкова, А.А. Самылин –
[Электрон. ресурс]. – 2005 /
Код доступа – http://www.ccr.ru/print.php?id=5975
21. Bollmark L. Seasonal dynamics and effects of nitrogen
supple rate on nitrogen and carbohydrate reserves in cutting–derived Salix
viminalis plants / L. Bollmark, L. Sennerby-Forsse, T. Ericsson // Can. J. Forest Res. – 1999. – 29,
№ 1. – P. 85 – 94.
22. Dawson W.M. Clonal selection in willow (Salix) grown as short rotation
coppice for energy production / W.M. Dawson, A.R. McCracken // Ann. Appl. Biol. – 1998. – 132, Suppl. –
P. 56–57.
23. McCracken A.R. Interaction
of willow (Salix) clones growing in mixtures // Ann. Appl. Biol. – 1998. – 132,
Suppl. – P. 54–55.
Подготовка и публикация
web-версии:
Афонин Алексей
Алексеевич
Доктор с.-х. наук,
профессор Брянского государственного университета
Зав. лабораторией
популяционной цитогенетики НИИ ФиПИ БГУ
e-mail: afonin.salix@gmail.com
главная страница сайта
ИВЫ: ТЕОРЕТИЧЕСКАЯ И ПРИКЛАДНАЯ САЛИКОЛОГИЯ http://afonin-59-salix.narod.ru
главная страница сайта
ОБЩАЯ И ТЕОРЕТИЧЕСКАЯ БИОЛОГИЯ http://afonin-59-bio.narod.ru