МЕТОДЫ АНАЛИЗА
МОРФОДИНАМИКИ ОДНОЛЕТНИХ ПОБЕГОВ ИВ (Salix)
А. А. Афонин, Брянский государственный университет, Россия
Главная страница сайта Ивы
(Salix, Willows, Weiden) <http://www.afonin-59-salix.narod.ru/index.html>
E-mail: <mailto:afonin.salix@gmail.com>
Исправленная и дополненная HTML-версия ранее опубликованных статей:
Афонин, А.А.
Метамерная изменчивость листьев ивы пепельной // Наука и образование –
возрождению сельского хозяйства России в XXI веке: Междунар. науч.-практ. и учебно-метод. конф. –
Брянск: Брянская ГСХА, 2000. – С. 70–72.
Афонин А.А. Ивы как
объект для изучения биологического разнообразия // Вестник БГУ. – 2003. – №1. –
С. 113–118.
Афонин А.А. Морфология
однолетних побегов ив // Вестник БГУ: Естественные и точные науки. – 2004. – №
4. – Брянск: БГУ, 2004. – С. 13–15.
Афонин А.А. Морфодинамика однолетних побегов ив // Лесопользование,
экология и охрана лесов: фундаментальные и прикладные аспекты. Материалы Междунар. науч.-практ. конф. – Томск: STT,
2005. – С. 223–224.
Изучение закономерностей изменчивости побегов древесных растений лежит в основе многих популяционно-морфометрических исследований (Магомедмирзаев, 1990). Для ив (род Salix) данная проблема наиболее актуальна, поскольку используемым продуктом этих растений, в основном, являются побеги (Левицкий, 1965).
Для создания устойчивых высокопродуктивных генетически гетерогенных насаждений ив используются клоносмеси, или поликлональные композиции – определенным образом подобранные сочетания саженцев, принадлежащих к разным сортам–клонам. В клоносмесях повышается общая устойчивость насаждений к экзогенным факторам, снижается вероятность распространения новых физиологических рас патогенов. При этом каждый клон должен обладать характеристиками, которые отсутствуют у других клонов (McCracken 1998).
На наш взгляд, важнейшей такой характеристикой является ритм развития побега.
Еще в 1917 г. Д’Арси Томпсон, основываясь на работах Гофмейстера и Бозе, обратил внимание на циклический характер развития растений (цит. по: Магомедмирзаев, 1990). В нашей стране первопроходцем в области изучения онтогенетической изменчивости был, несомненно, Н.П. Кренке (1940), проанализировавший одновершинные и двухвершинные возрастные кривые, отражающие жизнеспособность и мощность последовательно образующихся органов. К настоящему времени описаны одновершинные кривые изменения длины междоузлия вдоль побега для нескольких видов тимьяна (Берко, Козий, 1993b), михелии (Михалевская, 1993), ивы трехтычинковой (Михалевская, 1995), а также поузловая изменчивость размеров почек и листьев ивы розмаринолистной (Гетманец, 1994). Для расчета теоретически ожидаемых возрастных кривых предлагалось использовать кривые Пирсона (Кренке, 1940), методы гармонического анализа, основанные на вычислении коэффициентов полинома Фурье (Берко, Козий, 1993а). Таким образом, существует множество методик для описания ритмов развития побегов.
В качестве объектов исследования нами использовались выборки из природных популяций массовых древесных и кустарниковых видов ив БЛМ (система рода дана по А.К. Скворцову (1968), названия видов – по С.К. Черепанову (1995); далее для обозначения видов использованы латинские названия, а при сокращении – две первые латинские буквы): пятитычинковая (S. pentandra L. – pe), трехтычинковая (S. triandra L. – tr), ломкая (S. fragilis L. – fr), белая (S. alba L. – al), мирзинолистная (S. myrsinifolia Salisb. – my), козья (S. caprea L. – ca), пепельная (S. cinerea L. – ci), корзиночная (S. viminalis L. – vi), шерстистопобеговая (S. dasyclados Wimm. – da), остролистная (S. acutifolia Willd. – ac).
Для анализа морфодинамики однолетних побегов нами была разработана и использована следующая методика.
Для получения исходных данных по побегам с каждой особи определенного вида ив срезаются завершившие рост неразветвленные ростовые побеги (ауксибласты), показанные на рисунке.
Для фиксации первичных результатов описания побегов используются бланки. Указывается номер пробного участка, его географическое положение, ландшафтная принадлежность, эдафо-фитоценотическая характеристика. Фиксируется возрастное состояние особи (ювенильное, имматурное, виргинильное, раннее генеративное, зрелое генеративное, позднее генеративное, субсенильное, сенильное), наличие вторичных побегов, наличие повреждений, морфозов и терат. Объем материала определяется виталитетом особи, ее возрастным состоянием и количеством побегов, пригодных для описания; желательно с одной особи срезать не менее 10 побегов.
Все множество особей делится на три типа:
– с однородными побегами формирования;
– с явно неоднородными побегами формирования, образующими две-три группы побегов;
– с побегами, различающимися по внешнему виду и образующими непрерывный ряд.
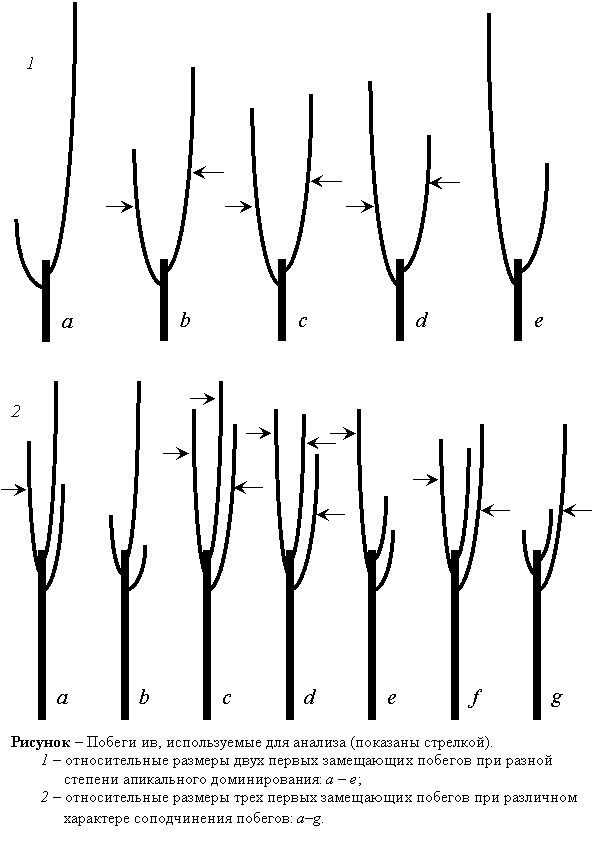
За структурно-функциональную единицу побега принимается метамер – узел с листом, пазушной почкой и нижележащим междоузлием (Магомедмирзаев, 1990). Для каждого метамера с порядковым номером t определяются следующие параметры (в мм): длина междоузлия I, длина черешка L, ширина листовой пластинки Х, длина листовой пластинки Y.
Для описания метамерной (порядковой, поузловой) изменчивости – зависимости отдельных параметров побега (I, L, X, Y) от порядкового номера метамера (узла) t используются морфодинамические ряды, графическим отражением которых являются морфодинамические кривые.
[Прилагательное «морфодинамический» использовалось ранее для обозначения особых точек (морфодинамических центров), концентрирующих энергию и воздействующих на вещество, собирающих его вокруг себя и становящихся видимыми и морфологически наблюдаемыми (Mawrodiadi, 1928, цит. по: Лепешинская, 1952). Мы вкладываем в этот термин иной смысл, и рассматриваем морфодинамический ряд (и его графическое отражение – морфодинамическую кривую) как ось креода – устойчивую информационно-энергетическую траекторию онтогенеза (морфогенеза), сформировавшуюся в результате канализирующего отбора (Waddington, 1940, цит. по: Уоддингтон, 1970).]
Эмпирические ряды динамики рассматриваются как случайные функции, например:
X (t) = φ (t) + U (t),
Y (t) = ψ (t) + V (t),
…………………….
где φ (t), ψ (t) – математические ожидания параметров Х, Y при заданных значениях порядкового номера метамера t («средние внутрикронные кривые»), а U(t), V(t) – случайные величины (Гмурман, 1999).
Для аппроксимации математических ожиданий случайных функций используются полиномы n-ой степени, например:
X = φ (t) = an · t n + ... + a1 · t + a0, Y = ψ (t) = bn · t n + ... + b1 · t + b0,
где n – целые числа (степень полинома), t – порядковый номер метамера, a1–n и b1–n – эмпирические коэффициенты, а0 и b0 – свободные члены уравнения (Львовский, 1982). Свободные члены (а0, b0,…) приравнивались к 0, тогда при t=0 значение любого параметра также равно 0.
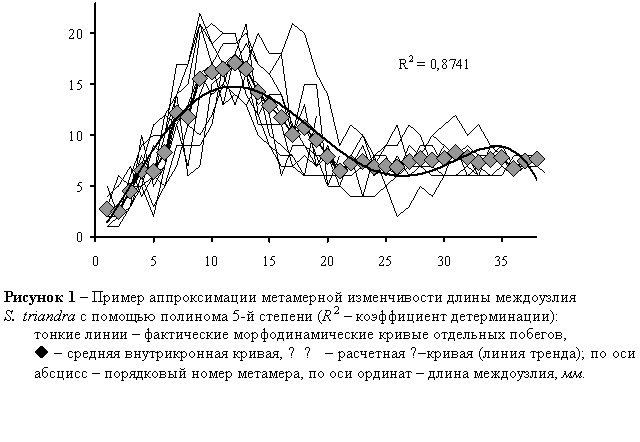
Количественная адекватность уравнений регрессии проверяется с помощью F-критерия Фишера (Львовский, 1982); качественная адекватность – с помощью коэффициентов аппроксимации R2. Степень полинома повышается до тех пор, пока прирост R2 не становился менее 0,1. Практически для достижения таких значений R2 оказывается достаточно пятой степени полинома. Тогда аппроксимирующие уравнения регрессии записываются следующим образом:
X=a5·t5+a4·t4+a3·t3+a2·t2+a1·t,
Y=b5·t5+b4·t4+b3·t3+b2·t2+b1·t,
………………………………………
Графики полученных уравнений регрессии (математических ожиданий случайных функций, расчетных морфодинамических кривых) мы условно назвали a–кривыми (например, X–a–кривая отражает метамерную изменчивость ширины пластинки, Y–a–кривая – метамерную изменчивость длины пластинки и т.д.).
Алгоритм построения a–кривых приведен в Приложении 1.
При сравнении a-кривых они рассматриваются как отражения гармонических (точнее, квазипериодических) колебаний со случайной амплитудой, случайной длиной волны и случайной фазой, описываемых уравнением:
X=А·е–δt·sin(ωt+φ)+b,
где X – значение признака в момент t (в данном случае t – порядковый номер метамера); А – мультипликативная компонента синусоиды, отражающая размах колебаний; (ωt+φ) – фаза гармонического колебания (ω – циклическая частота, φ – начальная фаза (–2π<φ<2π)); δ – коэффициент затухания, b – смещение по амплитуде (аддитивная компонента синусоиды, отражающая среднее значение признака); в свою очередь, ω=2πλ, где λ – длина волны (на побеге величине λ соответствует число метамеров между двумя смежными максимумами или минимумами).
При интерпретации полученных уравнений развивающийся побег рассматривалась как автоколебательная система, преобразующая внешние воздействия в периодические затухающие колебания и стремящаяся к стационарности с сохранением возможности смены режимов развития. На этом основании для качественного сравнения морфодинамических кривых (как эмпирических, так и расчетных) использовался понятийный аппарат гармонического анализа: смещение по фазе φ, по длине волны λ и по амплитуде (определяемое мультипликативной компонентой А и аддитивной компонентой b), затухание колебаний, переменная амплитуда и переменная длина волны (Натансон, 2001).
Для описания совместной метамерной изменчивости пар признаков используются параметрически заданные G–кривые на фазовой плоскости. Каждая G–кривая задается двумя уравнениями α–кривых (например, XY–G–кривая задается X- и Y–α–кривыми). Для качественного описания G-кривых используются термины «аттракторы», «особые точки», «предельные циклы» а также термины из области математического анализа (Натансон, 2001). Алгоритм построения G–кривых приведен в Приложении 2.
Для привязки локальных экстремумов α–кривых и особых точек G–кривые к определенным периодам вегетации используются прямые измерения длины вегетирующего побега (h, ±1 см) и диаметра несущего (прошлогоднего) побега на расстоянии 20 см от его основания (D20, ±0,1 мм) с интервалом 5…10 дней.
На основании анализа морфодинамики однолетних побегов нами были получены следующие результаты.
Метамерная изменчивость отдельных параметров побегов – закономерное изменение I, L, X, Y от основания побегов к вершине – носит периодический характер. Эмпирическая зависимость I, L, X, Y от порядкового номера метамера t адекватно описывается морфодинамическими α–кривыми – в большинстве случаев коэффициенты детерминации R2~0,9 (рисунок 1).
Максимальным разнообразием α–кривых характеризуется S.cinerea (до 10 типов), меньшим – S. myrsinifolia, S. triandra, S. alba, S. caprea, S. viminalis, S. dasyclados (5…8 типов), еще меньшим – S.pentandra, S.fragilis, S. acutifolia (2…4 типа).
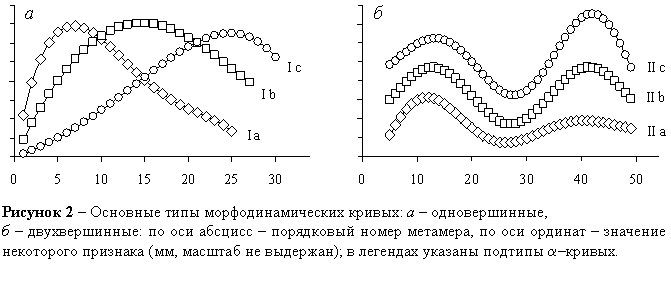
Периодический характер метамерной изменчивости I, L, X, Y обусловлен неравномерным развитием побегов в течение вегетационного сезона. Поэтому все многообразие a–кривых сводится к двум основным типам: одновершинным (с одной волной роста побега) и двухвершинным (с двумя волнами роста, разделенными периодом относительного покоя) (рисунок 2).
Одновершинные a–кривые делятся на три подтипа (рисунок 2а):
- Ia – абсолютный максимум в левой части a–кривой: наиболее интенсивное развитие побегов происходит в конце весны и в первой половине лета;
- Ib – абсолютный максимум вблизи средней части a–кривой: интенсивное развитие – в середине лета;
- Ic – абсолютный максимум в правой части a–кривой: интенсивное развитие – во второй половине лета.
Двухвершинные a–кривые также делятся на три подтипа (рисунок 2б):
- IIa – наиболее интенсивное развитие – в конце весны и в первой половине лета;
- IIb – весенне-летний и позднелетний периоды развития примерно равны по интенсивности;
- IIс–наиболее интенсивное развитие – во второй половине лета.
Разные типы и подтипы α–кривых характеризуются различной частотой. У всех изученных видов обнаружены одновершинные a–кривые I типа: I–a–кривые S. pentandra, S.triandra, S.caprea S.dasyclados и S.acutifolia, L–a–кривые S.pentandra, S.fragilis, S.caprea, S.cinerea, S.viminalis, S.acutifolia, X–a–кривые S.pentandra, S.fragilis, S.alba, S.viminalis, Y–a–кривые S.pentandra, S.fragilis, S.viminalis, S.dasyclados, S.acutifolia.Чаще встречается подтип Ia, реже – подтипы Ib и Ic. Некоторые подтипы одновершинных a–кривых характеризуются определенной видоспецифичностью, например, для S.triandra более характерны a–кривые с тенденцией к уменьшению I, X и Y к вершине побега, для S.alba – с тенденцией к увеличению Y. Двухвершинные α–кривые II типа с локальным минимумом выявляются на достаточно длинных побегах (свыше 20 узлов): I–a–кривые S.fragilis, S.alba, S.myrsinifolia, S.cinerea, S.viminalis, L–a–кривые S.triandra, S.alba, S.myrsinifolia, S.dasyclados, X–a–кривые S.myrsinifolia, S.caprea, S.cinerea, S.dasyclados, S.acutifolia, Y–a–кривые S.myrsinifolia, S.caprea, S.cinerea. Среди a–кривых II типа подтипы IIa, IIb, IIc встречаются с примерно одинаковой частотой. Не выявлены I–a–кривые II типа у S.pentandra, S.caprea, S.acutifolia, L–a–кривые у S.fragilis, S.viminalis, S.acutifolia, X–a–кривые у S.fragilis, S.alba, Y–a–кривые у S.fragilis.
При сравнении внутрикронных и индивидуальных a–кривых выявлено три типа различий между ними, которые описываются в терминах теории колебаний.
I Внутрикронные a–кривые сходны по конфигурации, незначительно различаясь по амплитуде А и длине волны λ. Тогда внутрикронные a–кривые характеризуют особь в целом, т.е. являются индивидуальными. В свою очередь, индивидуальные a-кривые отчетливо различаются по конфигурации (относятся к определенному типу или подтипу), что позволило выделить ритмотипы – группы особей одного вида, различающиеся по ритмам развития побегов. Сильнее индивидуальность a–кривых выражена у S.triandra, S.alba, S.myrsinifolia, S.caprea, S.cinerea, S.viminalis, S.dasyclados; слабее – у S.pentandra и S.acutifolia.
II Внутрикронные a–кривые существенно различаются по конфигурации (относятся к разным типам или подтипам), однако такая ситуация встречается редко: у отдельных особей S.fragilis (по I–a–кривым), S.cinerea (по I–, Х– и Y–a–кривым), S.viminalis (по Х–a–кривым).
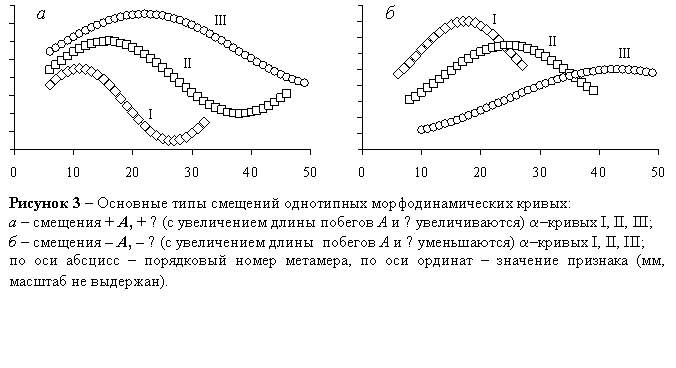
III Побеги разных групп разных особей содержат разное число узлов; тогда сходные по конфигурации a–кривые смещаются относительно друг друга по А и λ, и возникает проблема смещений. Выявлены следующие типы смещений: положительные [+] (бóльшими значениями А и/или λ характеризуются более длинные побеги), отрицательные [–] (бóльшими значениями А и/или λ характеризуются относительно короткие побеги), нейтральные [0] (А не зависит от длины побегов). Очевидно, внутрикронные смещения α–кривых обусловлены влиянием негенетических факторов, индивидуальные – влиянием и генетических, и негенетических факторов; при исключении негенетических факторов доказывается влияние генетических факторов. Основные типы смещений показаны на рисунке 3.
В системах внутрикронных и индивидуальных a–кривых выявляются морфодинамические аттракторы – области пересечения трех и более α–кривых: I–α–кривых S. fragilis, S. alba, S. myrsinifolia, S. cinerea, S. viminalis, S.dasyclados, L–α–кривых S. alba, S. viminalis, S. dasyclados, X–α–кривых S. pentandra, S. alba, S. cinerea, S. dasyclados, S. acutifolia, Y–α–кривых S. pentandra, S. triandra, S. myrsinifolia, S. acutifolia. Пример аттрактора в области пересечения индивидуальных X–a–кривых показан на рисунке 4.
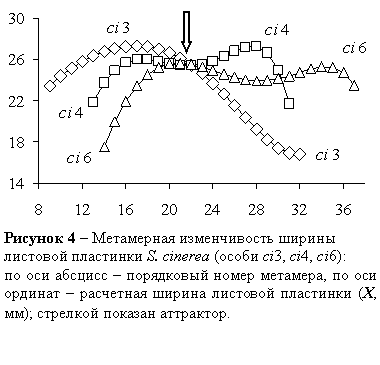
В некоторых случаях в пределах
одного вида наблюдается наложение отрезков двух и более α–кривых: I–α–кривых S.cinerea, S. acutifolia, L–α–кривых S.triandra, S. dasyclados; X–α–кривых S. pentandra, S.triandra, S. fragilis, S. myrsinifolia, S. dasyclados; Y–α–кривых S. pentandra, S. triandra, S. alba, S. acutifolia. В ряде случаев
совпадающие α–кривые завершаются точками бифуркации. У S. viminalis, S. dasyclados L–α–кривые образуют
разветвленно-сетевую систему. Таким образом, при наличии внутрикронных смещений
для презентации особи должна использоваться система a–кривых.
Сравнительный анализ взаиморасположения множества a–кривых показал, что влияние индивидуальных особенностей на положение a-кривых особенно велико у S. cinerea, в меньшей степени – у S. pentandra, S. triandra, S. alba, S. myrsinifolia, S. caprea. Для этих видов разнообразие ритмов развития отдельных параметров побегов рекомендуется для оценки биоразнообразия и прогноза успеха в селекции. Для других видов (S. fragilis, S.viminalis, S. dasyclados, S.acutifolia) эффективная селекция по метамерной изменчивости параметров побегов маловероятна. В целом наиболее эффективна оценка биоразнообразия и соответствующей эффективности селекционного процесса по метамерной изменчивости L (особенно у S. pentandra, меньше у S. myrsinifolia, S. caprea), менее эффективна – по метамерной изменчивости I (лишь у S. alba, S. cinerea), малоэффективна – по метамерной изменчивости X (допустима у S. caprea, S. cinerea); оценка по метамерной изменчивости Y возможна лишь у S. triandra (допустима у S. alba, S.myrsinifolia, S. cinerea). Вместе с тем, размеры листовой пластинки – это важный признак и с биологической точки зрения, и с хозяйственной (определяет продуктивность), и селекционной (облегчает визуальную оценку материала и отбор). Однако для анализа изменчивости X и Y требуются иные подходы, в частности, выявление закономерностей совместной изменчивости этих параметров с помощью параметрически заданных XY–G–кривых.
Параметрически заданные XY–G–кривые представлены несколькими типами с различной встречаемостью у разных видов (таблица 2).
Таблица 2 – Типы XY–G–кривых у разных видов ив
|
Типы XY–G–кривых |
Виды ив |
|||||||||
|
ca |
pe |
al |
da |
tr |
my |
ci |
vi |
ac |
fr |
|
|
Параболические |
– |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
|
С незамкнутыми эллипсами |
+ |
+ |
+ |
+ |
+ |
+ |
– |
– |
– |
– |
|
С замкнутыми эллипсами |
– |
– |
– |
– |
+ |
+ |
+ |
– |
– |
– |
|
N– и W–образные |
– |
– |
– |
+ |
+ |
+ |
+ |
+ |
+ |
– |
|
Обратные |
+ |
– |
+ |
– |
– |
– |
– |
+ |
– |
– |
|
Нестандартные |
– |
+ |
– |
– |
+ |
+ |
+ |
+ |
– |
+ |
|
Число стандартных типов |
2 |
2 |
3 |
3 |
4 |
4 |
3 |
3 |
2 |
1 |
Некоторые типы XY–G–кривых показаны на рисунке 5.
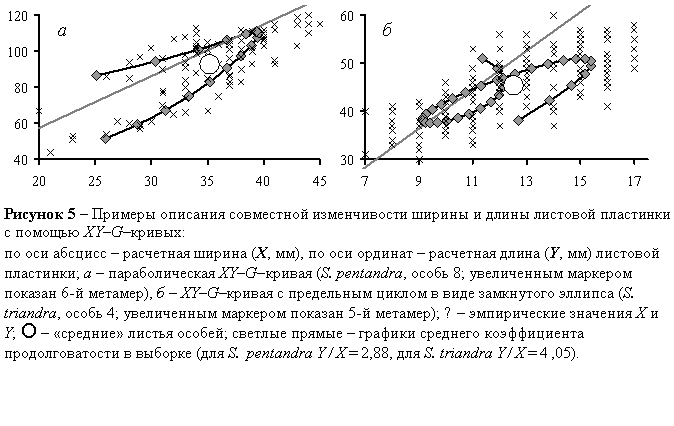
Параболические XY–G–кривые (рисунок 5а) имеют форму более или менее симметричной наклонной параболы, вершина которой лежит в области максимальных значений X и Y, а ось симметрии более или менее параллельна прямой, соответствующей среднему коэффициенту продолговатости Y/X. Обычно эти XY–G–кривые – гладкие, несамопересекающиеся, одновершинные, без особых точек. Однако в некоторых случаях парабола становится островершинной V-образной; такие кусочно-гладкие XY–G–кривые выявлены у S.pentandra, S.triandra, S.fragilis, S.alba, S.myrsinifolia, S.cinerea, S.viminalis, S.dasyclados, S.acutifolia.
XY–G–кривые с предельными циклами (рисунок 5б) завершаются предельными циклами в виде незамкнутых или полностью замкнутых эллипсов, длинные оси которых ориентированы вдоль прямых Y/X. Незамкнутыми эллипсами завершаются несамопересекающиеся XY–G–кривые (S.pentandra, S.triandra, S.alba, S. myrsinifolia, S.caprea, S.dasyclados), а полностью замкнутыми эллипсами – самопересекающиеся XY–G–кривые (S.triandra, S.myrsinifolia, S.cinerea).
Достаточно длинные побеги S.myrsinifolia, S.cinerea, S.viminalis, S.dasyclados, S.acutifolia характеризуются XY–G–кривыми с реверсией размеров листовой пластинки: N- и W-образными. В типичных случаях эти XY–G–кривые – гладкие, несамопересекающиеся, без особых точек; лишь в частных случаях выявляется самопересечение, появление острых вершин и предельных циклов.
Обратные XY–G–кривые наблюдаются в том случае, если у основания побегов листья более продолговатые, чем у вершины. Характерны для S.caprea, иногда обнаруживаются у S.alba, S.viminalis.
XY–G–кривые позволяют выявить больший уровень биоразнообразия, чем стандартные статистические показатели – M и σ2 (на рисунках 5а и 5b видно, что средняя величина может быть фиктивным показателем). Это обеспечивает более точную оценку материала при селекции, повышает эффективность селекционного процесса. У большинства видов (кроме S.fragilis, S.dasyclados, S.acutifolia) XY–G–кривые позволяют точнее разграничивать узколистные и широколистные особи. Индивидуальные различия по конфигурации XY–G–кривых сильнее всего выражены у S.triandra, в меньшей степени у S.fragilis, S.alba, S.myrsinifolia, S.cinerea. В то же время у S.pentandra, S.caprea, S.cinerea, S.viminalis, S.dasyclados, S.acutifolia общая конфигурация XY–G–кривых не может служить отличительным признаком особи. У этих видов межиндивидуальные различия между XY–G–кривыми могут быть меньше, чем межпобеговые – в результате XY–G–кривые разного типа характеризуют не особи, а группы побегов, и для презентации особей необходимо использовать системы внутрикронных XY–G–кривых.
Анализ морфодинамики основных параметров однолетних побегов ив показал, что использование a- и XY–G–кривых для оценки биоразнообразия и эффективности селекции ив наиболее эффективно для S. triandra; очевидно, природные популяции этого вида характеризуются высоким уровнем биоразнообразия, и при селекции легко выделяется множество отличимых клонов. Сложные подходы, основанные на сочетании анализа распределений и морфодинамики побегов, необходимы при работе с S. cinerea, в меньшей степени – S. pentandra, S.myrsinifolia. Некоторые виды – S.alba, S.caprea – нуждаются в дальнейшем изучении (в частности, в культуре). И наконец, для ряда видов – S. fragilis, S.viminalis, S. dasyclados, S.acutifolia – анализ изменчивости параметров однолетних побегов не рекомендуется для оценки биоразнообразия; у этих видов эффективная селекция по морфометрическим признакам маловероятна.
В то же время каждая морфодинамическая кривая представляет собой усредненный результат реализации генетической программы морфогенеза побегов (ГПМ) в определенных условиях генотипической и внешней среды. Иначе говоря, в генотипе особи запрограммирована устойчивая информационно-энергетическая траектория морфогенеза. Существование двух и более ГПМ в генотипе особи фенотипически проявляется в виде систем внутрикронных морфодинамических кривых, что приводит к формированию нескольких морфофункциональных групп побегов и обеспечивает гомеорез – устойчивость онтогенеза особи. Наличие множества программ морфогенеза в популяции обеспечивает гетерогенность насаждений по ритмам развития побегов, что позволяет более полно использовать ресурсы среды и повышает устойчивость к воздействию неблагоприятных факторов.
Главная
страница сайта Ивы (Salix, Willows, Weiden) <http://www.afonin-59-salix.narod.ru/index.html>
Библиография
Афонин, А.А. Метамерная изменчивость листьев ивы трехтычинковой / А.А. Афонин, Е.Н. Самошкин // Лесоведение. – 2006. – № 2. – С. 5–10.
Берко И.Н., Козий Б.И. Методика гармонического анализа морфологических параметров метамерного строения монокарпических побегов растений / Жизненные формы: онтогенез и структура. – М.: Прометей, 1993. – С. 145–150.
Берко И.Н., Козий Б.И. Моделирование метамерного строения монокарпических побегов и его значение для систематики рода Thymus L. (Lamiaceae) / Жизненные формы: онтогенез и структура. – М.: Прометей, 1993. – С. 140–145.
Гетманец И.А. Морфологические особенности побегов и жизненные формы Salix rosmarinifolia L., Salix repens L. / Успехи экологической морфологии растений и ее влияние на смежные науки. – М.: Прометей, 1994. – С. 20–21.
Гмурман В.Е. Теория вероятностей и математическая статистика. – М.: Высш. шк., 1999. – 479 с.
Левицкий И.И. Ива и ее использование. - М.: Лесн. пром-сть. 1965. – 98с.
Лепешинская О.Б. Развитие жизненных процессов в доклеточном периоде. – Изд-во АН СССР: М.: 1952. – 202 с.
Львовский Е.Н. Статистические методы построения эмпирических формул. – М.: Высш. шк., 1982. – 224 с.
Магомедмирзаев М.М. Введение в количественную морфогенетику. – М.: Наука, 1990. – 229 с.
Михалевская О.Б. Ритм роста и структура побега у видов Magnolia L. и Michelia L. (Magnoliaceae) / Жизненные формы: онтогенез и структура. – М.: Прометей, 1993. – С. 135–139.
Михалевская О.Б. Структура и развитие боковых силлептических побегов у древесных растений и представление об элементарном побеге / Актуальные вопросы экологической морфологии растений. – М.: Прометей, 1995. – С. 70–76.
Кренке Н.П. Основные положения теории циклического старения и омоложения растений в онтогенезе (Теория возрастной цикличности) / Теория циклического старения и омоложения растений. – М.: Огиз–Сельхозгиз, 1940. – С.5-31.
Натансон И.П. Краткий курс высшей математики. – СПб.: Лань, 2001. – 736 с.
Уоддингтон К.Х. Основные биологические концепции / На пути к теоретической биологии. – М.: Мир, 1970. – Ч.1. – Пролегомены. – С.11–38.
McCracken A. R. Interaction of willow (Salix)
clones growing in mixtures (Взаимодействие клонов ивы при
выращивании в смесях) // Ann. Appl. Biol. – 1998. – 132, Suppl. – P.
54 – 55. – РЖ ВИНИТИ. – 2000 – №2.
Главная
страница сайта Ивы (Salix, Willows, Weiden) <http://www.afonin-59-salix.narod.ru/index.html>
Приложение П1. Алгоритм
построения α–кривых
1. Составляется таблица, в ячейках которой указывается значение параметра (например, Y, мм) для определенного метамера (t) каждого побега, подобная таблице 1.1.
Таблица П.1.1 – Форма для описания метамерной (порядковой, поузловой) изменчивости (исходные данные)
|
Порядковый номер метамера, t |
Номера побегов |
|||||
|
1 |
2 |
3 |
4 |
5 |
6 |
|
|
1 |
– |
– |
– |
– |
– |
– |
|
2 |
– |
– |
– |
– |
– |
– |
|
3 |
– |
– |
– |
– |
– |
– |
|
4 |
46 |
– |
– |
– |
52 |
– |
|
5 |
69 |
– |
– |
47 |
70 |
52 |
|
6 |
84 |
– |
51 |
63 |
80 |
63 |
|
7 |
88 |
– |
70 |
66 |
77 |
66 |
|
8 |
95 |
92 |
73 |
72 |
85 |
70 |
|
9 |
100 |
|
67 |
80 |
87 |
60 |
|
10 |
103 |
86 |
57 |
88 |
77 |
67 |
|
11 |
94 |
104 |
71 |
80 |
63 |
65 |
|
12 |
84 |
83 |
65 |
85 |
80 |
55 |
|
13 |
91 |
83 |
62 |
85 |
67 |
59 |
|
14 |
94 |
96 |
66 |
84 |
65 |
56 |
|
15 |
88 |
92 |
– |
79 |
71 |
50 |
|
16 |
91 |
85 |
– |
65 |
43 |
50 |
|
17 |
92 |
92 |
– |
– |
– |
49 |
|
18 |
82 |
84 |
– |
– |
– |
– |
|
19 |
– |
77 |
– |
– |
– |
– |
|
20 |
– |
36 |
– |
– |
– |
– |
|
21 |
– |
65 |
– |
– |
– |
– |
|
22 |
– |
60 |
– |
– |
– |
– |
|
23 |
– |
59 |
– |
– |
– |
– |
|
24 |
– |
40 |
– |
– |
– |
– |
2. На основании подобной таблицы строится график зависимости значения параметра от порядкового номера метамера (рисунок П.1.1).
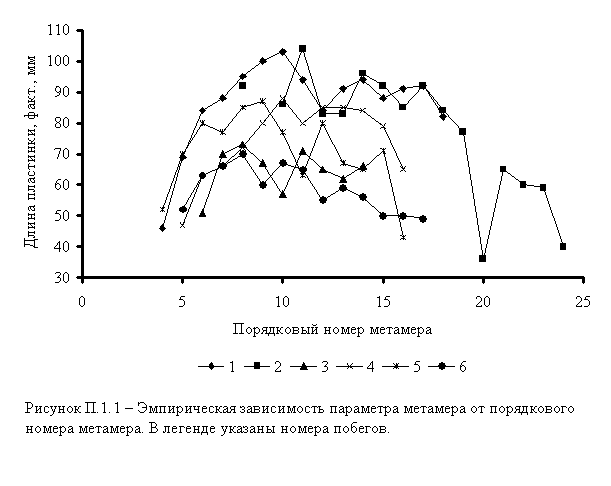
Визуальный анализ рисунка П.2.2.1.1 показывает, что по характеру метамерной изменчивости побеги 1 и 2 резко отличаются от остальных побегов особи. Поэтому исходный массив данных разбиваем на два подмассива, на две группы побегов (количество групп побегов может быть различным у разных особей – от 1 до 3). Для каждой группы побегов составляем таблицу, подобную таблице 1.2, причем в таблицу вводятся столбцы N и «Среднее».
Таблица П.1.2 – Преобразованная форма таблицы для описания метамерной изменчивости (упорядоченные исходные данные)
|
Порядковый номер метамера, t |
Среднее |
N |
Номера побегов |
|||
|
3 |
4 |
5 |
6 |
|||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
|
4 |
52,00 |
1 |
|
|
52 |
|
|
5 |
56,33 |
3 |
|
47 |
70 |
52 |
|
6 |
64,25 |
4 |
51 |
63 |
80 |
63 |
|
7 |
69,75 |
4 |
70 |
66 |
77 |
66 |
|
8 |
75,00 |
4 |
73 |
72 |
85 |
70 |
|
9 |
73,50 |
4 |
67 |
80 |
87 |
60 |
|
10 |
72,25 |
4 |
57 |
88 |
77 |
67 |
|
11 |
69,75 |
4 |
71 |
80 |
63 |
65 |
|
12 |
71,25 |
4 |
65 |
85 |
80 |
55 |
|
13 |
68,25 |
4 |
62 |
85 |
67 |
59 |
|
14 |
67,75 |
4 |
66 |
84 |
65 |
56 |
|
15 |
66,67 |
3 |
|
79 |
71 |
50 |
|
16 |
52,67 |
3 |
|
65 |
43 |
50 |
Примечание. N – количество побегов с пригодными для описания метамерами; в столбцах приведены значения параметра; среднее рассчитывается как частное от деления суммы параметров в строке на величину N, например, для 5-го метамера среднее значение параметра равно (47+70+52)/3=56,33.
3. На основании таблицы П.1.2 строится эмпирическая «средняя внутрикронная кривая» – график зависимости среднего значения признака от порядкового номера метамера (рисунок П.1.2).
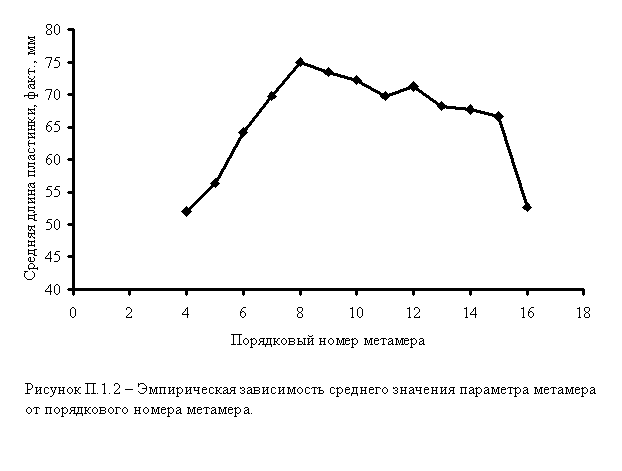
4. Дальнейшая задача состоит в аппроксимации эмпирической последовательности. Коэффициенты полиномов 5-й степени вычисляются с помощью встроенных ресурсов Excel (рисунок П.1.3).
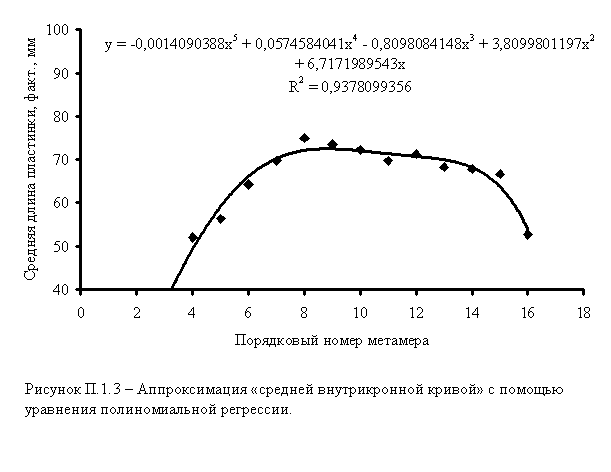
Полученные полиномы (и соответствующие им кривые) представляют собой
интерполяционные эмпирико-статистические модели, отражающие общие закономерности
внутрипобеговой изменчивости отдельных параметров побега.
Приложение П.2. Алгоритм построения параметрически
заданных G-кривых
На основании рассчитанных морфодинамических рядов (математических ожиданий случайных функций) каждому расчетному значению одного параметра можно поставить расчетное значение других параметров (таблица 2).
Таблица П.2 – Расчетные значения параметров побега
|
Порядковый номер метамера, t |
Расчетные значения
параметров, мм |
|||
|
I |
L |
X |
Y |
|
|
8 |
10,72 |
9,56 |
17,81 |
54,88 |
|
9 |
13,13 |
11,08 |
21,72 |
62,23 |
|
10 |
15,60 |
12,51 |
25,48 |
69,08 |
|
11 |
18,02 |
13,81 |
28,89 |
75,41 |
|
12 |
20,25 |
14,91 |
31,74 |
81,19 |
|
13 |
22,17 |
15,75 |
33,86 |
86,39 |
|
14 |
23,66 |
16,30 |
35,10 |
90,97 |
|
15 |
24,63 |
16,53 |
35,35 |
94,83 |
|
16 |
24,99 |
16,43 |
34,59 |
97,84 |
|
17 |
24,69 |
16,00 |
32,84 |
99,78 |
|
18 |
23,73 |
15,28 |
30,22 |
100,35 |
|
19 |
22,13 |
14,33 |
26,94 |
99,14 |
|
20 |
19,98 |
13,25 |
23,32 |
95,64 |
|
21 |
17,43 |
12,16 |
19,81 |
89,16 |
|
22 |
14,67 |
11,24 |
16,98 |
78,89 |
|
23 |
12,01 |
10,70 |
15,57 |
|
|
24 |
9,81 |
10,79 |
16,44 |
|
|
25 |
8,51 |
11,82 |
|
|
|
26 |
8,68 |
|
|
|
При парном анализе расчетному значению одного параметра (например, Xt) можно поставить в соответствие расчетное значение другого параметра (например, Yt) и описать совместную внутрикронную изменчивость признаков на фазовой плоскости X–Y. Полученная кривая и есть параметрически заданная G–кривая.
Ниже приведены пример полученных α–кривых и соответствующей XY–G-кривой одной из особей ивы пятитычинковой.
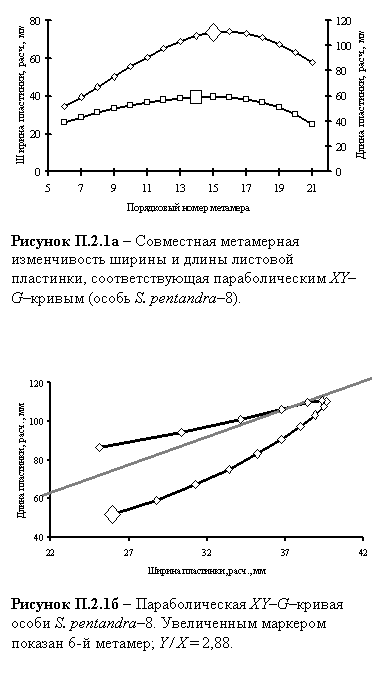
Главная
страница сайта Ивы (Salix, Willows, Weiden) <http://www.afonin-59-salix.narod.ru/index.html>