Морфодинамика однолетних побегов длиннолистных форм
ивы трехтычинковой
Афонин Алексей
Алексеевич,
Анищенко Лидия
Николаевна,
Борздыко Елена
Васильевна
Брянский
государственный университет им. акад. И.Г. Петровского,
Лаборатория
популяционной цитогенетики НИИ ФиПИ
г. Брянск
Оригинал статьи:
© Афонин А.А.
Морфодинамика однолетних побегов длиннолистных форм ивы трехтычинковой / А.А.
Афонин, Л.Н. Анищенко, Е.В. Борздыко // Наука и современность – 2010: Сб.
материалов V междунар. научно-практ.
конфер. – Ч.1. – Новосибирск: Изд-во НГТУ, 2010. – С. 30–34.
Ива
трехтычинковая (Salix triandra L.) – массовый вид
кустарниковых ив средней полосы России, перспективный для создания насаждений
комплексного назначения. Из природных популяций S. triandra Брянского
лесного массива выделены длиннолистные формы. Для
выявления различий в морфодинамике однолетних побегов использованы
параметрически заданные XY-G-кривые. Показано, что разные
длиннолистные особи характеризуются разными ритмами развития. Выявленные дендроритмотипы рекомендованы для селекции S. triandra на
отличимость, а также для создания поликлональных насаждений в различных
эдафо-фитоценотических условиях.
Ивы
(Salix L.) – широко известные листопадные
фанерофиты – занимают первое место в дендрофлоре средней полосы европейской
части России по видовому разнообразию [Скворцов, 1968]. Одним из наиболее
распространенных видов Salix
является ива трехтычинковая (S.
triandra L.). Этот вид традиционно используется как источник
белого и зеленого прута, который применяется для плетения корзин, домашней
утвари, изгородей. Легкость размножения черенками делает иву трехтычинковую
незаменимой для создания противоэрозионных насаждений [Левицкий, 1965]. Ива
трехтычинковая перспективна как источник биомассы для нужд биоэнергетики: её
продуктивность сопоставима с продуктивностью ивы корзиночной (S. viminalis L.) [Логинова, 2010] – популярным
продуцентом биотоплива «второго поколения» [Егоров, 2005; Цивенкова, Самылин,
2005; Dawson,
McCracken,
1998], но, в отличие от указанного типичного аллювиально-пойменного вида, ива
трехтычинковая успешно произрастает и в менее благоприятных
эдафо-фитоценотических условиях: на сравнительно переувлажненных, недостаточно
аэрируемых дерново-подзолисто-глеевых, иловато-болотных, торфяно-глеевых и
маломощных торфяных почвах [Афонин, 2003, 2009b].
Ива
трехтычинковая характеризуется ареалом евро-азиатского типа,
бореально-средиземноморского подтипа (Средняя и Атлантическая Европа, Южная Скандинавия,
Средиземноморье, Северная Африка, Кавказ, Малая Азия, Иран, Западная и
Восточная Сибирь, Дальний Восток, Монголия, Япония, Китай). Вследствие
географической изменчивости описаны географические расы видового и подвидового
ранга: S.
nipponica Fr. et Sav. 1876 = ssp.
nipponica Skvorts.; S.Bornmuellerii Haussk. 1890 = ssp. bornmuellerii
Skvorts.;
S.
Kinashii
Levl.
1909; S.
Medwedewii
Dode,
1908; S.
hamatidens
Levl.
1909. Описаны межвидовые гибриды с участием ивы трехтычинковой: S.viminalis × S. triandra
(S.
undulata
– волнистолистная); S.
triandra
× S.
viminalis
(S.
molissima);
S.
purpurea
× S.
undulata
(гибрид
Сукачева); S.
triandra
× S.purpurea
(ива
американская); S.
triandra
× S.
alba;
S.
cinerea
× S.
triandra.
Кроме того, ива трехтычинковая характеризуется высоким уровнем
внутрипопуляционного полиморфизма [Валягина-Малютина, 2004], что дало основание
выделить разновидности: var.
concolor
Hort.
= f.
viridis Spenn. (листья с двух сторон зеленые); var. discolor (S.
amygdalina L.) = ssp. amygdalina = f. glaucophylla Ser. (листья снизу беловато-сизые); var. vulgaris Wimm. (листья ланцетные); var. elliptica Toeppf. (листья эллиптические); var. latifolia Schatz. (Toeppf.)
(листья широкоэллиптические); var.
angustifolia Ser. (листья узкие) [Скворцов, 1968].
На
основании многолетних наблюдений за изменчивостью ивы трехтычинковой на
территории Брянского лесного массива (БЛМ) [Афонин, 2003] нами было
установлено, что типичные экземпляры S. triandra характеризуются
продолговато-эллиптическими листьями длиной 55...80 (30...100) мм при ширине
13...20 (6...27) мм. Однако морфология листа может сильно изменяться вдоль
побега вследствие метамерной (порядковой, поузловой) изменчивости: в
проксимальной части побегов листья эллиптические, яйцевидные, по мере
продвижения к вершине становятся продолговато-эллиптическими,
продолговато-ланцетовидными. Метамерная изменчивость затрудняет идентификацию
внутривидовых форм, что препятствует оценке генетического потенциала природных
популяций и дальнейшей селекции на отличимость. Однако в 1999 г. на территории
БЛМ в одной из природных ценопопуляций S. triandra нами была выявлена и
описана узко-длиннолистная форма [Афонин, Самошкин, 2006]. В 2009…2010 гг. был
осуществлен целенаправленный поиск узко-длиннолистных форм в
различных ценопопуляциях S.
triandra
на
территории Снежетьско-Деснянского низменного (долинно-речного) ландшафта БЛМ. В 2009 г. в качестве
материала использовались весенние (безлистные) черенки, из которых в салицетуме
Брянского государственного университета (БГУ) были выращены особи черенкового
происхождения. В 2010 г. материал собирался в природных популяциях в виде
облиственных побегов.
В
конце вегетационного периода 2010 г. с каждой особи, обладающей типичными
признаками вида, срезались неразветвленные ростовые побеги (ауксибласты). За
структурно-функциональную единицу побега был принят метамер – узел с листом,
пазушной почкой и нижележащим междоузлием [Магомедмирзаев, 1990]. Для каждого
метамера с порядковым номером t с
точностью ±0,5 мм измерялись ширина (Х) и длина (Y) листовой пластинки.
Для создания баз данных и обработки результатов: вычисления средних значений X
и
Y (M±m)
и коэффициента продолговатости Y/X – использовался процессор Excel.
Метамерная
(порядковая, поузловая) зависимость ширины и длины листовой пластинки от
порядкового номера метамера (узла) t описывалась с помощью морфодинамических рядов, графическим
отражением которых являются морфодинамические
α-кривые [Афонин, 2000]. Эмпирические α-кривые рассматривались как реализации случайных функций:
X (t)
= u
(t)
+ U
(t),
Y (t)
= v
(t)
+ V
(t),
где u(t) и v(t) –
генетически детерминированные математические ожидания Х и Y при
заданных значениях порядкового номера метамера t, а U(t) и V(t) –
случайные величины [Гмурман, 1999], зависящие от комплекса парагенетических
факторов.
Для
аппроксимации математических ожиданий случайных функций использовались полиномы
5-й степени:
X = a5·t5
+ a4·t4 + a3·t3
+ a2·t2 + a1·t + a0,
Y = b5·t5
+ b4·t4 + b3·t3
+ b2·t2 + b1·t + b0,
где а0…5 и b0…5
– эмпирические коэффициенты.
Полученные
уравнения регрессии использовались для построения параметрически заданных XY-G-кривых на фазовой плоскости XY.
Особые области XY-G-кривых на фазовой плоскости представляют
собой морфодинамические аттракторы [Афонин, 2009а].
Для
окончательного анализа были отобраны три особи с заметно удлиненными листьями:
– tr-993 – асс. Alno-Salicetum
cinereae,
координаты 34.507728 в.д., 53.201025 с.ш., побеги для исследования срезаны
4.08.1999; Xср. = 14,3±0,14 мм, Yср. = 82,0±0,97 мм, Y/X=5,7;
– tr-091 – черенки
заготовлены 20.04.2009 в асс. Salicetum triandro-viminalis, координаты 34,382311 в.д.,
53,255248 с.ш.; затем укоренены и выращены в коллекционном салицетуме БГУ, побеги
для исследования срезаны 10.08.2010; Xср. = 15,3±0,40 мм, Yср. = 89,5±3,38 мм,
Y/X=5,8;
– tr-102 – асс. Alno-Salicetum cinereae, координаты 34,494578 в.д.,
53,203577 с.ш., побеги для исследования срезаны 4.08.1999; Xср. = 16,3±0,23 мм,
Yср. = 103,0±2,46
мм, Y/X=6,3.
В качестве
объекта сравнения использовалась ранее описанная [Афонин, Самошкин, 2006] особь
tr-st (tr998, асс. Salicetum albae, координаты 34.280276 в.д., 53.19283 с.ш.;
побеги для исследования срезаны 27.07.1999) с типичной морфологией листьев и
стандартной U-образной XY-G-кривой; Xср. = 12,4±0,21 мм, Yср. = 45,3±0,51 мм,
Y/X=3,6.
Фазовые портреты
побегов названных особей показаны на рисунке.
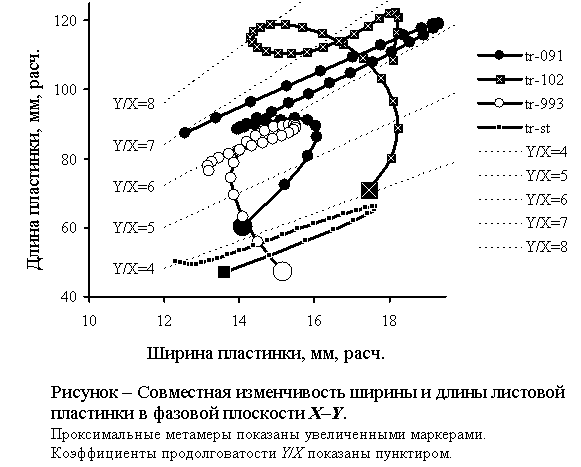
Сравнительный
анализ полученных параметрически заданных XY-G-кривых показал следующее.
1)
Морфодинамика побегов особи tr-091 описывается W-образной XY-G-кривой с двумя морфодинамическими
аттракторами в виде «сгущения» маркеров: в области X=14…15
мм, Y=89…93 мм и области X≈19
мм, Y=113…119 мм. В обоих случаях Y/X≈6,5.
2.
Особь tr-993 характеризуется
самопересекающейся T-образной
XY-G-кривой, с одним морфодинамическим
аттрактором: X=14…16 мм, Y=81…90
мм, Y/X≈6,0.
3.
XY-G-кривая особи tr-102
имеет сложную конфигурацию: она также самопересекающаяся, но аттрактор в этом
случае определяется не коэффициентом продолговатости Y/X,
а петлей, описываемой вокруг воображаемого центра с координатами X=15
мм, Y=115 мм, Y/X=7…8.
Таким
образом, выделенные формы ивы трехтычинковой отличаются от типичной особи tr-st и друг от друга по длине листовой
пластинки, что дает основание назвать их длиннолистными (f. longifolia).
В то же время, эти формы различаются по ритмам развития побегов, что позволяет
использовать их в селекции на отличимость, а также для создания генетически
гетерогенных поликлональных насаждений [Афонин, 2009с].
Список
литературы
1.
Афонин А.А. Ивы Брянского лесного
массива. – В 2-х частях. – Брянск: БГУ, 2003. – 237 с.
2.
Афонин А.А. Метамерная изменчивость
листьев ивы пепельной // Наука и образование – возрождению сельского хозяйства
России в XXI
веке: Междунар. науч.-практ. и учебно-метод. конф. – Брянск: Брянская гос.
с.-х. академия, 2000. – С. 70–72.
3.
Афонин А.А. Метамерная изменчивость
листьев ивы трехтычинковой / А.А. Афонин, Е.Н. Самошкин // Лесоведение. – 2006.
– № 2. – С. 5–10.
4.
Афонин А.А. Методы анализа морфодинамики
однолетних побегов ив (Salix).
[Электрон. ресурс]. – 2009а /
Код доступа: http://afonin-59-salix.narod.ru/060825_dynamica.htm
5.
Афонин А.А. Популяционная биология ив Среднего
Подесенья // Синтетическая теория эволюции: состояние, проблемы, перспективы. –
Материалы Междунар. конфер., посвящ. 200-летию со дня рожд. Ч. Дарвина и
150-летию выхода книги «Происхождение видов путем естественного отбора…». –
Луганск: Элтон-2, 2009b.
– С. 53-55.
6.
Афонин А.А. Селекция ив на разнообразие
ритмов развития // Фактори
експериментальноï еволюцiï организмiв: збiрник наукових праць (Материалы V
междунар. научн. конфер. «Факторы экспериментальной эволюции организмов»). –
Т.6. – Киïв:
Логос, 2009c.
– С. 259–263.
7.
Валягина-Малютина Е.Т. Ивы европейской
части России. – М.: КМК, 2004. – 217 с.
8.
Гмурман В.Е. Теория вероятностей и
математическая статистика. – М.: Высш. шк., 1999. – 479 с.
9.
Егоров А.И. Ивовая энергетика [Электрон. ресурс]. – 2005 / Код доступа: http://hepd.pnpi.spb.ru/ioc/ioc/line051112/n5.htm
10.
Левицкий И.И. Ива и ее использование. –
М.: Лесн. пром-сть, 1965. – 98 с.
11.
Логинова
Л.А. Продуктивность и энергетический потенциал ивовых ценозов на примере
Воронежской области. – Автореф. дис… канд. биол. наук. – Воронеж, 2010.
12.
Магомедмирзаев М.М. Введение в
количественную морфогенетику. – М.: Наука, 1990. – 229 с.
13.
Скворцов А.К. Ивы СССР. – М.: Наука,
1968. – 262 с.
14.
Цивенкова Н.М. Леса – неисчерпаемые
энергоресурсы России / Н.М. Цивенкова, А.А. Самылин [Электрон.
ресурс]. – 2005 / Код доступа: http://www.ccr.ru/print.php?id=5975
15.
Dawson
W.M. Clonal selection in willow (Salix) grown as short rotation coppice for
energy production / W.M. Dawson, A.R. McCracken // Ann. Appl. Biol. – 1998. –
132, Suppl. – P. 56–57.
Подготовка и
публикация web-версии:
Афонин Алексей
Алексеевич
Доктор с.-х.
наук, профессор Брянского государственного университета
Зав.
лабораторией популяционной цитогенетики НИИ ФиПИ БГУ
e-mail: afonin.salix@gmail.com
главная страница
сайта ОБЩАЯ И ТЕОРЕТИЧЕСКАЯ БИОЛОГИЯ http://afonin-59-bio.narod.ru
главная страница
сайта ИВЫ6 ОБЩАЯ И ТЕОРЕТИЧЕСКАЯ САЛИКОЛОГИЯ http://afonin-59-salix.narod.ru
последнее обновление страницы 12.03.2012