СОХРАНЕНИЕ И ПРИУМНОЖЕНИЕ УРОВНЯ БИОЛОГИЧЕСКОГО
РАЗНООБРАЗИЯ
В КВАЗИНАТИВНЫХ ЭКОСИСТЕМАХ
Тезисы доклада на Всероссийской конференции-семинаре «Гетерогенность биологических систем и вариабельность их реакций на действие факторов окружающей среды» (Институт биологии Коми НЦ УрО РАН, г. Сыктывкар)
Выходные данные:
Афонин, А.А. Сохранение и приумножение уровня биологического разнообразия в квазинативных экосистемах / А.А. Афонин, Л.И. Булавинцева // Биологические эффекты малых доз ионизирующей радиации и радиоактивное загрязнение среды: Материалы Междунар. конф. (Сыктывкар, Республика Коми, Россия, 28 сентября – 1 октября 2009 г.). – Сыктывкар: Коми НЦ УрО РАН, 2009. – С. 391–393.
В современном мире, когда воздействие человека на биосферу приобрело глобальный характер, на Земле практически не сохранились природные (нативные) экосистемы, эволюционирующие по естественным законам. Основным компонентом биосферы становятся квазинативные экосистемы: внешне похожие на естественные, но сложившиеся под влиянием антропогенных факторов. Типичные примеры квазинативных экосистем: заброшенные сельхозугодья, вырубки и вторичные леса, экосистемы нарушенных местообитаний (пустыри, полосы отчуждения). При снижении антропогенного стресса в таких экосистемах начинаются вторично-сукцессионные процессы, которые (по крайней мере, теоретически) могут привести к восстановлению их естественной, эволюционно устойчивой структуры. Общепризнано, что эволюционная устойчивость экосистем тесно связана с уровнем биологического разнообразия. Эволюционная устойчивость – это способность экосистемы сохранять существующий уровень биоразнообразия на протяжении длительного числа поколений при отсутствии катастроф. В то же время, оптимальный уровень биоразнообразия обеспечивает устойчивость экосистем к воздействию внешних факторов.
На протяжении последних десятков лет в общественно-научном сознании сложилось мнение, что основу биоразнообразия составляет альфа-разнообразие (число видов в экосистеме). Отсюда следует вывод о том, что монокультура не может быть устойчивой. Однако фактически наиболее устойчивыми являются экосистемы, в которых альфа-разнообразие сравнительно невелико. Например, в экосистемах равнинных европейских лесов каждый род относительно крупных организмов, как правило, представлен одним видом (реже двумя или немногими видами). Это легко объясняется принципом конкурентного исключения («один вид – одна ниша»). Но тогда получается, что проблема сохранения биоразнообразия как фактора устойчивости носит искусственный характер. В то же время, известны многовидовые генетически гетерогенные экосистемы, отличающиеся высокой устойчивостью к воздействию внешних факторов. Примером таких экосистем служат ивняки, в которых средообразующими видами-эдификаторами являются ивы (Salix L.) – RS-стратеги, известные высоким уровнем видового и внутривидового разнообразия. В современных условиях можно наблюдать ивняки, в различной мере подверженные воздействию антропогенных факторов: от почти ненарушенных многовидовых ивняков на болотах до одновидовых (моно-, би- и поликлональных) плантаций.
Для анализа последствий конкуренции в подобных многовидовых генетически гетерогенных биологических системах нами были разработаны эколого-генетические детерминантные имитационные модели, основанные на допущениях Лотки-Вольтерра и Харди-Вайнберга. В качестве выходной функции отклика рассматривалась переменная NT – численность особей данного вида или генотипа в зрелом генеративном состоянии в некотором поколении Т. Функция отклика NT зависит от ряда входных переменных: в первую очередь, от исходной численности популяции каждого вида N0 и от мальтузианского параметра r, который мы рассматривали, как число гамет, оставляемых одной особью данного вида или гено/фенотипа. Рассматривались значения N0 порядка 102 (чтобы минимизировать влияние случайных факторов) и значения r≥2 (что обеспечивает самовоспроизведение генотипа). В отсутствие конкуренции N0 и r характеризуют кривизну экспоненциальной функции увеличения численности группы (популяции, генотипа) в идеальных условиях. В этом случае r является постоянной величиной – фундаментальным аутэкологическим параметром.
Для описания генетической структуры панмиктической популяции (а популяции видов Salix можно считать таковыми за счет большого радиуса эффективного распространения семян) использовался показатель p(A) – частота аллеля (гаплотипа) «дикого типа» А, который не изменяется качественно на протяжении всего рассматриваемого эволюционного периода. Множество изоаллелей (изогаплотипов) а1…an рассматривалось как общий альтернативный аллель (гаплотип) а с частотой q. Рассматривались стартовые значения p(A) в интервале 0,01…0,99. Распределение генотипов среди особей нулевого возраста (семян, проростков) рассчитывалось по алгоритму Харди-Вайнберга: P(AA)=p2(AA), H(Aa)=2pq(Aa), Q(aa)=q2(aa); P+H+Q=1.
Конкурентные отношения между генотипами в популяциях каждого вида и биотически-симпатрическими популяциями разных видов описывались с помощи матрицы коэффициентов конкуренции α – показателей, отражающих интенсивность конкурентных отношений между генотипами в одной популяции или между разными взаимодействующими популяциями. Коэффициенты aii…ajj отражают напряженность внутригрупповой конкуренции (в зависимости от контекста, в пределах одного гено/фенотипа или одной популяции). Матрицы коэффициентов aij отражают интенсивность конкуренции между группами i и j (в зависимости от контекста, конкуренции между гено/фенотипами одной популяции или конкуренции между разными популяциями). Пример такой матрицы приведен в таблице 1. Множитель (1–a11·N01–a12·N02…–a1k·N0k), где k – число конкурирующих групп (гено/фенотипов или популяций биотически-сипатрических видов), определяет синэкологическое изменение мальтузианского параметра r в уравнениях Лотки-Вольтерра. Если межгрупповые взаимодействия симметричны, то коэффициенты межгрупповой конкуренции равны, например, a12=a21 (это идеальная ситуация). Если же межгрупповые взаимодействия несимметричны, то эти коэффициенты, разумеется, неравны: a12≠a21 (эта ситуация ближе к реальной). Если N0≈102, то a≈10–4. При составлении матриц коэффициентов aij мы исходили из дарвиновского представления о том, что напряженность внутригрупповой конференции выше, чем межгрупповой; чем больше экологические различия между группами, тем ниже напряженность межгрупповой конкуренции. Пример такой матрицы для диаллельной системы приведен в таблице 2.
Варьируя и комбинируя значения N0, r, p(A), α, нам удалось проанализировать сотни эволюционных сценариев. Выводы, полученные для разных виртуальных популяций, не противоречат друг другу. В качестве примера рассмотрим результаты анализа динамики генофонда одной из виртуальной популяций: N0=400, r=3 (независимо от генотипа), p0(A)=0,900; значения α взяты из таблицы 2.
В
данной виртуальной популяции действует частотно-зависимый отбор: фактическая
приспособленность гено/фенотипов зависит от их частоты. Тогда с возрастанием
абсолютной частоты каждого гено/фенотипа его приспособленность снижается за счет
возрастания внутригрупповой конкуренции (рис. 1).
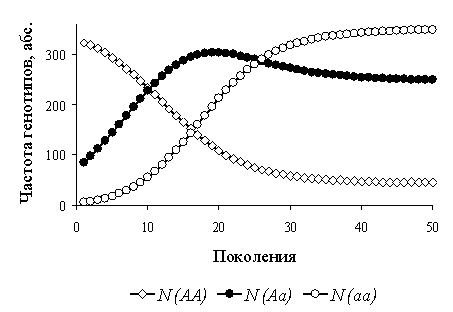
Рисунок 1 – Динамика абсолютных частот гено/фенотипов в виртуальной популяции
С достижением численности популяции, максимально возможной в данных условиях (полное использование доступных ресурсов), устанавливается равновесная частота p(A), которая определяется матрицей коэффициентов конкуренции. Внешне это выглядит как смена движущего отбора (Δp(A)≠0) стабилизирующим отбором на разнообразие: Δp(A)=0, 0>p(A)>1 (рис. 2).
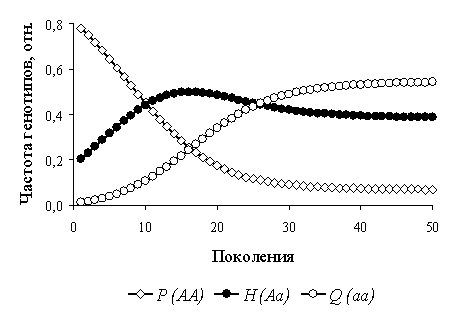
Рисунок 2 – Динамика относительных частот гено/фенотипов в виртуальной популяции
В этом случае монокультуры оказываются генетически гетерогенными и эволюционно устойчивыми.
Формирование эволюционно устойчивого многовидового сообщества (например, из видов А и В) возможно только при абсолютном равенстве межвидовых коэффициентов конкуренции (aАВ=aВА), при этом коэффициенты внутривидовой конкуренции могут быть любыми (но больше, чем коэффициенты межвидовой конкуренции). Однако фактически этого не бывает (aАВ≠aВА), поэтому устойчивость многовидовых сообществ (точнее, синузий) носит относительный характер, и сукцессия завершается формированием монодоминантного сообщества. В итоге «побеждает» тот вид, который в данных условиях более конкурентоспособен (в предельном случае аменсализма aАВ=0, а aВА>0).
Сформулированные положения носят дискуссионный характер, поэтому авторы воздерживаются от формулировки окончательных выводов и рекомендаций. Однако результаты, полученные при анализе предложенных моделей, позволяют иначе взглянуть на проблему сохранения и приумножения биологического разнообразия.
Одновидовая «монокультура» с высоким уровнем генетического разнообразия оказывается эволюционно более устойчивой, чем «поликультура» с высоким видовым, но низким внутривидовым разнообразием. Поэтому при проектировании искусственных эволюционно устойчивых экосистем следует избегать избыточно высокого уровня видового разнообразия, поскольку увеличение числа видов приводит к снижению уровня внутривидового (внутрипопуляционного) разнообразия за счет уменьшения численности популяций отдельных видов.
При анализе структуры существующих квазинативных экосистем следует обращать внимание не столько на разнообразие видов (альфа-разнообразие), сколько на разнообразие аллелофонда популяционных компонентов. Для сохранения имеющегося уровня внутривидового генетического разнообразия необходимо поддерживать максимально высокую численность популяций (сотни особей в генеративном состоянии), а для повышения изначально низкого уровня этого разнообразия – вводить новые источники изменчивости.
В прикладном аспекте, генетическое разнообразие оценивается как разнообразие гено/фенотипов. Однако высокий уровень генотипического разнообразия (например, средняя гетерозиготность на полиморфный локус) может наблюдаться при обедненной структуре аллелофонда, а именно, при сниженной доле полиморфных генов (на практике – генов, у которых частота редкого аллеля не выше 0,1). С точки зрения эволюционной устойчивости нас интересует именно доля таких генов, подавляющее большинство которых не проявляется на морфологическом уровне. Поэтому при введении отличимых гено/фенотипов (применительно к ивам, это маркировка по окраске покровов побегов, по морфологии листьев) новый материал следует брать из пространственно изолированных популяций.
Поскольку многовидовые сообщества эволюционно неустойчивы при постоянных коэффициентах конкуренции, то следует добиваться максимальной гетерогенности среды с тем, чтобы в каждом микроместообитании складывалась своя матрица этих коэффициентов. Этого легко добиться, если местообитание изначально неоднородно (например, поймы рек; здесь уровень альфа-разнообразия ив значительно выше, чем на водоразделах). Если же эдафотоп изначально гомогенен, то необходимо принимать меры для повышения уровня его мозаичности. Изменять рельеф и почвенный покров – дело неблагодарное и рискованное. Для создания первичной неоднородности местообитания легче изменить фитоценотическую среду путем создания областей сгущения–разрежения.
В настоящее время нами разрабатываются тригонально-центрированные схемы-модули размещения гено/фенотипов (клонов) разных видов. Эти модули принципиально отличаются от предложенных нами ранее гексагонально-центрированных схем размещения клонов, которые были разработаны для оптимизации конкурентных отношений в устойчивых поликлональных насаждениях без смены поколений. Такие гексагонально-центрированные (или просто гексагональные) модули воплощают идею «абсолютного равенства». Тригонально-центрированные модули предназначены для создания «изначального неравенства», которое должно обеспечить устойчивость насаждения на протяжении многих поколений. В основе тригонально-центрированных схем лежат две подсистемы из равносторонних конгруэнтных треугольников, смещенных относительно друг друга на 1/3 высоты этих треугольников. Эти подсистемы могут быть представлены как видами, близкими в эколого-таксономическом отношении (например, ива белая – ива трехтычинковая), так и далекими (например, ива белая – липа мелколистная). Поскольку в реальном эксперименте проследить эволюционную судьбу популяций древесных растений не предоставляется возможным, то предполагается оценивать внутривидовую и межвидовую конкурентоспособность по изменению ритмов развития побегов.
Ожидаемый результат – сохранение структуры насаждения на протяжении ряда поколений – может быть получен только через многие десятки лет после начала эксперимента. Однако анализ промежуточных результатов позволит уточнить теоретические основы управления квазинативными экосистемами, что станет еще одним шагом на пути к рациональному неистощительному природопользованию.
ЛИТЕРАТУРА
Афонин А.А. Ивы как объект для изучения биологического разнообразия // Вестник БГУ. Брянск: Брянский гос. ун-т, 2003. №1. С. 113–118.
Афонин А.А. Методологические принципы создания устойчивых высокопродуктивных насаждений ив (на примере автохтонных видов Salix Брянского лесного массива). Брянск: РИО Брянского гос. ун-та, 2005. 146 с.
Афонин А.А. Ивы [Электрон. ресурс] / http://afonin-59-salix.narod.ru
Булавинцева Л.И., Анищенко Л.Н., Зайцев Д.Н. Руководство к практическим занятиям по технологии и методике обучения биологии. Часть 2. Приложения к практическим занятиям. Брянск: РИО БГУ, 2008. 176 с.
Дарвин Ч. Происхождение видов путём естественного отбора. Пер. с 6 изд. (Лондон, 1872). / Отв. редактор А.Л. Тахтаджян. СПб: Наука, 2001. 568 с.
Динамика
популяционных генофондов при антропогенных воздействиях / Под
ред. Ю.П. Алтухова. М.: Наука, 2004. 619 с.
Милютин
Л.И. Генетико-эволюционные основы устойчивости лесных экосистем // Лесоведение.
– 2003. – №1. – С. 16–20.
Пианка Э. Эволюционная экология. М.: Мир, 1981. 400 с.
Примак Р.Б. Основы сохранения биоразнообразия. М.: НУМЦ, 2002. 256 с.
© Афонин Алексей Алексеевич
Доктор с.-х. наук, профессор Брянского государственного университета
Зав. лабораторией популяционной цитогенетики НИИ ФиПИ БГУ
e-mail: afonin.salix@gmail.com
главная страница сайта ОБЩАЯ И ТЕОРЕТИЧЕСКАЯ БИОЛОГИЯ http://afonin-59-bio.narod.ru
главная страница сайта Ивы:
Теоретическая и прикладная саликология http://www.afonin-59-salix.narod.ru
дополнительные web-ресурсы
http://afonin-59-salix.narod.ru
последнее обновление страницы 05.09.2012