РАСЩЕПЛЕНИЕ ПО МОРФОЛОГИЧЕСКИМ ПРИЗНАКАМ
ПРИ СЕМЕННОМ РАЗМНОЖЕНИИ ИВЫ ПЛАКУЧЕЙ
В ТРЕХ ПОКОЛЕНИЯХ
(25 августа 2006 г.)
А. А. Афонин, Брянский государственный университет, Россия
Главная страница сайта Ивы (Salix, Willows, Weiden) <http://www.afonin-59-salix.narod.ru/index.html>
E-mail: <mailto:afonin.salix@gmail.com>
Исправленная и дополненная HTML-версия ранее опубликованных материалов:
статья –
Афонин А.А. Расщепление по морфологическим признакам при семенном воспроизведении ивы плакучей // Наука и образование – возрождению сельского хозяйства России в XXI веке. – Междунар. научно-практ. и учебно-метод. конфер. – Брянск: Изд-во Брянской ГСХА, 2000. – С. 73–75.
монография –
Афонин А.А. Методы изучения разнообразия ив в коллекционных насаждениях. – Брянск: Изд-во Брянского государственного университета, 2002. – 67 с. – УДК 582.623+634.928.53+634.956.58+631.52
Плакучие ивы широко используются для озеленения населенных пунктов и закрепления берегов водоемов.
Под названием «плакучая ива» обычно объединяются различные плакучие формы (культивары cv – культивируемые разновидности) ивы белой (Salix alba L.): Salix alba f. pendula hort (L.), Salix alba f. vitellina pendula Rend. (плакучая крона с блестяще-желтыми побегами).
К плакучим ивам относится также ива вавилонская (S. babylonica); однако в России этот вид культивируется редко из-за низкой морозоустойчивости.
Гораздо чаще культивируются гибриды ивы вавилонской и белой, которые обладают сравнительно высокой морозоустойчивостью – S. solomonii Carriere (S. babylonica ´ S. alba) и S. alba var. tristis Ser. («ива печальная» – S. babylonica ´ S. alba var. vitellina).
Реже разводят гибриды ивы ломкой (S. fragilis) и вавилонской (S. babylonica), например, S. Í blanca Anders. (S. babylonica ´ S. fragilis).
Родословную некоторых клонов (культиваров) в настоящее время трудно установить.
В юго-западных регионах России часто выращивается клон гибридного происхождения S. babylonica ´ S. alba, который характеризуется следующими признаками.
По габитусу (внешнему облику) это быстрорастущее дерево с шатровидной или раскидистой плакучей кроной, образованной многочисленными тонкими разветвленными побегами длиной до 2 метров и более. Кора вегетирующих побегов зеленая, затем желтеющая.
Листья на изогнуто-закрученных желобчатых коротких черешках, мелко-железисто-пильчатые, узко яйцевидно-ланцетовидные, с клиновидным основанием и длиннозаостренной саблевидно изогнутой верхушкой; длина черешка 0,6...1,5 см, ширина пластинки 1...2 см при длине 8...15 см. Пластинка достигает максимальной ширины ближе к своему основанию.
В целом, листья кожистые, разноцветные: сверху темно-зеленые, снизу сизые. У основания пластинки с верхней стороны 1...2 пары мелких железок – сидячих или на ножках. Опушение листовой пластинки снизу и сверху шелковистое, опушение черешков и молодых стеблей щетинисто-бархатистое. Почки мелкие, узкие, прижатые; опушение щетинисто-бархатистое, переходящее на верхушке в шелковистое. Прилистники некрупные (длиной в несколько мм), сидячие, асимметричные узко ланцетовидные, рано опадающие.
Однолетний побег рассматриваемого клона условно можно разделить на три части. В базальной части побега происходит увеличение длины пластинки примерно от 40 до 100 мм при сохранении средней ширины 12...13 мм. В средней части побега длина пластинки медленно возрастает от 100 до 120 мм, а ширина от 13,5 до 15,5 мм с сохранением отношения длины к ширине 7,5:1...7,6:1. В апикальной части побега происходит стремительное уменьшение размеров пластинки с увеличением отношения длины к ширине до 10:1. Таким образом, базальные листья являются продолговато–эллиптическими, срединные – продолговатыми, а апикальные – крайне продолговатыми (мы предлагаем называть их extremum oblongum).
Сережки сходны с сережками ивы белой. Сережки андрогинные – содержат и пестичные, и тычиночные цветки. Пыльца часто деформирована; длина нормальных пыльцевых зерен 28,98±0,261 мкм (что сопоставимо с размерами пыльцевых зерен ив подрода Salix – пятитычинковой (S. pentandra), ломкой (S. fragilis), белой (S. alba)).
Случаи спонтанного или искусственного семенного возобновления данного культивара ивы плакучей до начала наших опытов нам неизвестны.
Расщепление в исходном поколении (F0– 1994 г.)
Для изучения расщепления по морфологическим признакам в исходном поколении при семенном возобновлении ивы плакучей нами использовались семена, собранные с особи, обладающей типичными признаками описанного культивара. Всхожесть семян – практически 100%-ная. Однако далее началась гибель сеянцев. Критическими периодами в летний период оказались стадия второго настоящего листа (гибель в конце первого месяца жизни) и стадия 5…8 настоящих листьев (гибель в конце лета). В зимний период погибли сеянцы, имеющие меньше 8 настоящих листьев, а в конце зимы – начале весны и более развитые сеянцы. Таким образом, в генотипе рассматриваемого клона имеются многочисленные нежелательные генетические факторы (аллели), приводящие к гибели значительной части потомков, что препятствует воспроизведению этого культивара самосевом. В итоге весной 1995 г. лишь 4 однолетние особи (cvvp1…4) были высажены в открытый грунт: на среднеокультуренной песчаной почве (В4) при высокой освещенности.
Сеянец cvvp1 к сентябрю 1999 г. (в 6 лет) представлял собой сильное одноствольное дерево с раскидистой полуплакучей кроной высотой > 6 м. С 1998 г. эта особь обильно цвела, в сережках только пестичные цветки. В настоящее время сеянец срублен, но от него выращены черенковые саженцы (раметы cvvp1r), которые показывают высокую устойчивость к заболеваниям и высокую продуктивность (годичный прирост в высоту > 2 м при диаметре двухлетних ветвей > 4 см).
Сеянец cvvp2 к сентябрю 1999 г. достиг высоты 4 м. По габитусу напоминал дикорастущую иву белую, однако сережки андрогинные – с пестичными и тычиночными цветками. В настоящее время сеянец срублен.
Сеянец cvvp3 – карликовый (< 1 м). Крона раскидистая, плакучая. Однолетние побеги обильно ветвились. Усиленный рост побегов – во второй половине лета. Сережки отсутствовали до 2000г. включительно. Таким образом, нежелательные аллели исходного клона могут проявиться и среди выживших потомков. В настоящее сеянец cvvp3 срублен.
Сеянец cvvp4 обладает наиболее высокими декоративными качествами. Это стройное дерево с ровным стволом, раскидистой полуплакучей кроной, тонкими краснеющими однолетними побегами, высотой более 3 м (1999 г.) – примерно в 2 раза меньше, чем у cvvp1. Цветет с 1998г., сережки только пестичные.
В целом клон cvvp1 может рекомендоваться для разведения как наиболее сильный, а клон cvvp4 – как среднерослый с красивыми побегами.
Расщепление в первом поколении (F1– 1999 г.)
В 1999 г. были собраны семена сеянцев cvvp1 и cvvp4, опыленных пыльцой сеянца cvvp2. Всхожесть семян с особи cvvp1 составила почти 100%, с особи cvvp4 примерно 50%. Основываясь на закономерностях классической генетики, можно предположить, что генотипы особей cvvp2 и cvvp4 содержат вредные аллели. В определенных сочетаниях эти аллели приводят к снижению общей продуктивности особей, а в других сочетаниях – к гибели зародышей. В начале 2000 г. особь cvvp2 была срублена, и весной вновь были собраны семена с особей cvvp1 и cvvp4. Все семена оказались невсхожими. Таким образом, подтверждается, что именно особь cvvp2 была донором пыльцы, и исключается возможность апомиксиса. Из потомства сеянца cvvp1 зимовку успешно перенесли 13 особей, а из потомства сеянца cvvp4 – только одна; таким образом, генотип сеянца cvvp4 действительно насыщен нежелательными аллелями.
На втором году жизни (в начале июня 2000 г.) сеянцы F1 были высажены в открытый грунт на коллекционном участке БГУ. В качестве контроля высажена рамета cvvp1r (1999г.), сохраняющая признаки материнского растения cvvp1.
В начале сентября 2001 г. была произведена предварительная оценка двухлетних сеянцев F1. Расщепление наблюдалось, в первую очередь, по габитусу. Часть сеянцев сформировала один прямой неразветвленный лидерный побег. У двух особей произошло раннее опадание листьев в базальной и средней части побега, и деревца приобрели пальмовидный облик. Другая часть сеянцев развивалась с образованием длинного лидера с обильным ветвлением в средней и верхней части кроны. И, наконец, часть сеянцев (в том числе, и у единственного потомка особи cvvp4) образовала множество побегов от основания лидерного побега. По окраске побегов выщепились цветовые формы. Большинство растений – с желто-зеленой окраской побегов (как у исходного клона); у части из них побеги краснеющие. Две особи имели светло-зеленые побеги без пигментации. У двух особей побеги зеленовато-коричневые (как у дикорастущей ивы белой). И, наконец, одна особь характеризовалась темно-ярко-красной окраской коры, стеблей и почек; листья интенсивно-темно-зеленые. По общей морфологии листья сеянцев F1 продолговато-ланцетовидные. У большинства сеянцев F1 побеги без прилистников, но у части особей на вершине побегов имелись мелкие полусердцевидные прилистники (fragilis–тип), а у части – мелкие шиловидные (alba–тип).
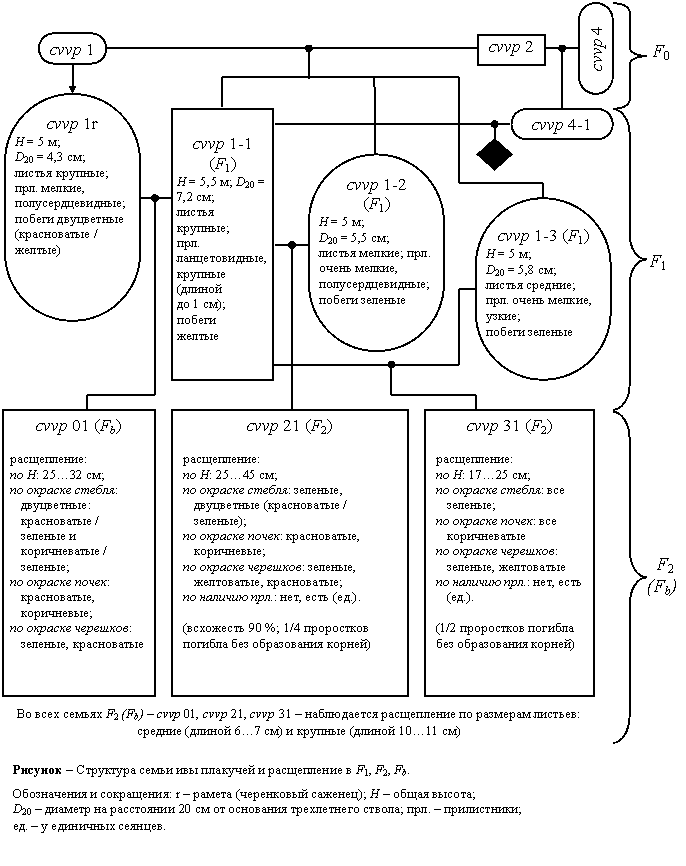
Из 13 потомков особи cvvp1 устойчивый прирост сохранился только у четырех; у остальных 9 темпы годичного прироста в высоту и по диаметру на протяжении 3…7 лет снижались (очевидно, в ходе онтогенеза проявились нежелательные аллели отцовской особи cvvp2). В течение 2002–2006 гг. самые слабые сеянцы были удалены. Сохранившиеся сильные сеянцы (cvvp1-1…4, cvvp4-1) – дважды обрезались на пень. В начале сентября 2005 г. (по двухлетним стволам на 6-летних корнях) было произведено их очередное описание по двухлетним стволам.
Максимальным приростом в высоту характеризуется сеянец cvvp1-1 (прирост за два года 4,5 м, в т.ч. 2,5 м за первый год). У других потомков особи cvvp1, её раметы cvvp1r и сеянца cvvp4-1 прирост за два года составил 3,5…4,2 м (в т.ч. за первый год 1,5…2,1 м). По диаметру двухлетних стволов также лидирует сеянец cvvp1-1 (D=5,0 см), далее идут сеянцы cvvp1-3 (D=4,6 см), cvvp1-2 (D=4,1 см) и рамета cvvp1r (D=4,2 см), что значительно превышает прирост в естественных насаждениях S. alba (Морозов, 1966). У сеянцев cvvp4-1 и cvvp1-4 показатели прироста в толщину заметно ниже (D=3,3 и 2,9 см). Объем двухлетней части ствола у сеянца cvvp1-1 превысил 2 дм3 (в пересчете на сухую древесину ~ 1 кг); у сеянцев cvvp1-3, cvvp1-2 и раметы cvvp1r – 1150…1380 см3 (в пересчете на сухую древесину ~530…630 г). При густоте посадки 10 тыс. шт./га это составит по объему ~ 6…10 м3/гаÍгод, по воздушно-сухой массе ~ 3…5 т/гаÍгод.
В трех поколениях отмечено морфологическое сходство между исходным клоном cvvp0, сеянцем cvvp1 и его потомком cvvp1-1: в первую очередь, по яично-желтой окраске зимних побегов и двулетних ветвей – таким образом, наблюдается прямое наследование признаков. В то же время, указанные особи различаются по морфологии сережек и листьев.
Обращает на себя сходство сеянцев cvvp1-2 и cvvp4-1: несмотря на то, что они происходят от разных материнских особей, однолетние побеги сходны по окраске покровов и черешков (буровато-красные), а также листовых пластинок (интенсивная темно-зеленая окраска). В данном случае наблюдается сцепление по окраске разных частей побега. Длина годичного прироста в первый год после обрезки примерно одинаковая (~2м), но диаметр двухлетних стволов у cvvp1-2 в 1,25 раза больше (4,1 см, H/D ~ 50 против 3,3 см, H/D ~ 60); то есть при равном годичном приросте в длину объем древесины у cvvp1-2 ~ в 1,5 раза больше. Таким образом, прирост по диаметру ствола в данном случае не зависит от окраски побегов и длины годичного прироста.
У большинства сеянцев (кроме cvvp1-4) крона полуплакучая. В целом наиболее мощные сеянцы – cvvp1-1 и cvvp1-3. Сеянец cvvp1-2 отличается прямым стволом, обильным ветвлением; сеянец cvvp1-4 – прямым стволом и яйцевидно-конической кроной. У перечисленных сеянцев высота трехлетних стволов на 7-летних корнях – 5…5,5 м.
Черенки сеянцев cvvp1-1 и cvvp1-2 хорошо укореняются, что дает возможность размножать полученные биотипы вегетативным путем. Черенки сеянца cvvp1-3 в открытом грунте укореняются плохо, т.е. высокая продуктивность данной особи сочетается с ослабленным ризогенезом.
Расщепление во втором поколении (F2 и Fb– 2006 г.)
У большинства сеянцев F1 нормальное цветение началось весной 2006г. (в 7 лет, на 2-летних стволах): у особей cvvp1-2, cvvp1-3, cvvp4-1 сережки пестичные, у cvvp1-1 – тычиночные.
Длина пыльцевых зерен сеянца cvvp1-1 – 32,26±0,220 мкм (в 1,11 раза больше, чем у исходного клона), что сопоставимо с размерами пыльцевых зерен ив подрода Vetrix – пепельной (S. cinerea), мирзинолистной (S. myrsinifolia), шерстистопобеговой (S. dasyclados).
Однодомных растений в F1 нет; таким образом, однодомность в данной семье наследуется как рецессивный признак.
Для изучения расщепления во втором поколении были произведены сибсовые скрещивания сеянца ♂cvvp1-1 с особями ♀cvvp1-2, ♀cvvp1-3, ♀cvvp4-1 и возвратное скрещивание ♀cvvp1r Í ♂cvvp1-1. В большинстве пар были получены нормальные семена, и только в паре ♀cvvp4-1 Í ♂cvvp1-1 (полусибсы) семена оказались мелкими, с почти нулевой всхожестью; единичные проростки (семядоли слабые, корни не образовывались) погибли в первые дни жизни. Среди однолетних сеянцев, полученных в результате успешных скрещиваний, наблюдались расщепления по окраске стеблей, черешков, жилок, почек; яркая окраска стеблей коррелирует с яркой окраской черешков. Большинство сеянцев с яркой окраской частей побега погибло в первые месяцы жизни. Особенности расщепления среди самых сильных выживших сеянцев в отдельных семьях показаны на схеме (рисунок).
Больше всего форм (в т.ч. с максимальной высотой однолетних сеянцев) выщепилось в потомстве cvvp1-2, меньше всего – в потомстве cvvp1-3, хотя эти особи и сходны между собой по окраске побегов. Это означает, что с точки зрения менделизма по цветовым признакам особь cvvp1-2 гетерозиготна в большей степени, а особь cvvp1-3 – в меньшей. В потомстве раметы cvvp1r стебли сеянцев двуцветные (зеленых нет); следовательно, генетические факторы зеленой окраски стебля отсутствуют у особей cvvp1r и cvvp1-1, но имеются у cvvp1-2 и cvvp1-3. Во всех семьях F2 (Fb) наблюдается расщепление по размерам листьев; следовательно, донор пыльцы ♂cvvp1-1 и ♂cvvp1r (крупнолистные) гетерозиготны по этому признаку. Максимальная высота сеянцев наблюдается в потомстве cvvp1-2, минимальная – в потомстве cvvp1-3. Это означает, что особи cvvp1-1 и cvvp1-2 гетерозиготны в наибольшей степени, особь cvvp1-3 – в меньшей, а рамета cvvp1r занимает промежуточное положение. В потомстве cvvp1-2 и особенно cvvp1-3 у части проростков не образовалась корни. Таким образом, особи cvvp1-1, cvvp1-2 и cvvp1-3 содержат вредные аллели, подавляющие ризогенез, причем у особи cvvp1-3 их больше (что подтверждается слабым корнеобразованием на черенках); у раметы cvvp1r эти аллели отсутствуют. В целом анализ расщеплений в F2 (Fb) показал, что высокая продуктивность сеянцев F1 определяется их гетерозиготностью. Для получения семян желательно использовать особи типа cvvp1-2, у которых достаточно высокая продуктивность сочетается с оптимальным содержанием нежелательных аллелей.
Таким образом, изученная форма ивы плакучей генетически неоднородна и может использоваться как исходный материал для выведения новых декоративных и высокопродуктивных устойчивых форм.
 |
Библиография
1 Анциферов Г.И. Ива. – М.: Лесная промышленность, 1984. – 101 с.
2 Валягина-Малютина Е.Т. Ивы европейской части России. – М.: КМК, 2004. – 217 с.
3 Левицкий И.И. Ива и ее использование. – М.: Лесн. пром-сть, 1965. – 98 с.
4 Морозов И.Р. Ивы СССР, их использование и применение в защитном лесоразведении. – М.–Л.: Гослесбумиздат, 1950. – 167 с.
5 Морозов И.Р. Определитель ив и их культура. – М.: Лесн. пром-сть, 1966. – 254 с.
6 Правдин Л.Ф. Ива, ее культура и использование. – М.: Изд-во АН СССР, 1952. – 168 с.
7 Рычин Ю.В. Древесно-кустарниковая флора. – М.: Просвещение, 1972. – 264 с.
8 Скворцов А.К. Сем. Salicaceae – Ивовые // Флора средней полосы европейской части СССР. – М.: Колос, 1964. – С. 184–199.
9 Скворцов А.К. Ивы СССР. – М.: Наука, 1968. – 262 с.
10 Скворцов А.К. Salicaceae Mirbel – Ивовые // Флора европейской части СССР. – Л.: Наука, 1981. – С. 10–33.
11 Старова Н.В. Селекция ивовых. – М.: Лесн. пром-сть, 1980. – 206 с.
12 Федоров Ал.А., Кирпичников М.Э., Артюшенко З.Т. Атлас по описательной морфологии высших растений. Лист. / Под общ. ред. П.А. Баранова. М.–Л.: Изд-во АН СССР, 1956. 303 с.
13 Царев, А. П. Селекция и репродукция лесных древесных пород / А. П. Царев, С. П. Погиба, В. В. Тренин. - М.: Логос, 2003 – 503 с.